
The Wrong Standard for Performance

Performance, as most athletes track it, is a record of what they can do at their best. The metric is the heaviest lift, the fastest split, the sharpest sparring round in recent memory. This framing is intuitive and it produces a clear number to aim at, but it measures the wrong thing. A fighter who has one sharp night and two flat ones in the same week has not performed well. He has performed unpredictably. A professional in his forties who trains at full intensity Tuesday and loses the back half of the week to recovery debt has not produced a training week. He has borrowed from the next five days.
What actually matters, particularly for athletes who carry significant professional and personal load alongside their training, is repeatability. Not the ceiling of output, but the floor: how consistently the system produces quality work across sessions, weeks, and training years without progressively degrading the biological infrastructure that makes output possible.
PEAK OUTPUT
What most athletes optimize for
The best effort, the heaviest lift, the fastest split. Measures the ceiling of performance at a single point in time. Produces a clear personal record with no information about repeatability.
DURABLE OUTPUT
What actually compounds
Consistent, repeatable performance across sessions, weeks, and training years. Measures the floor of performance — how reliably quality work is produced under load.
Durability is not toughness. Toughness is a psychological quality involving willingness to tolerate discomfort. Durability is a physiological state: the biological capacity for consistent, repeatable output without degrading the systems that generate it. The fitness-fatigue model provides the biological framework for making that distinction.
These two qualities are often conflated, and the confusion is costly. An athlete who is genuinely psychologically tough but not physiologically durable will push through warning signals, interpret fatigue as motivation failure, and accelerate the breakdown of the very systems he is trying to develop. Understanding where one ends and the other begins determines how to read what is happening in the body and what to do about it.
Durability is a physiological state: the biological capacity for consistent, repeatable output without degrading the systems that generate it.
An athlete who is genuinely psychologically tough but not physiologically durable will push through warning signals, interpret fatigue as motivation failure, and accelerate the breakdown of the very systems he is trying to develop.
The Fitness-Fatigue Model: How Training Either Builds or Erodes Performance
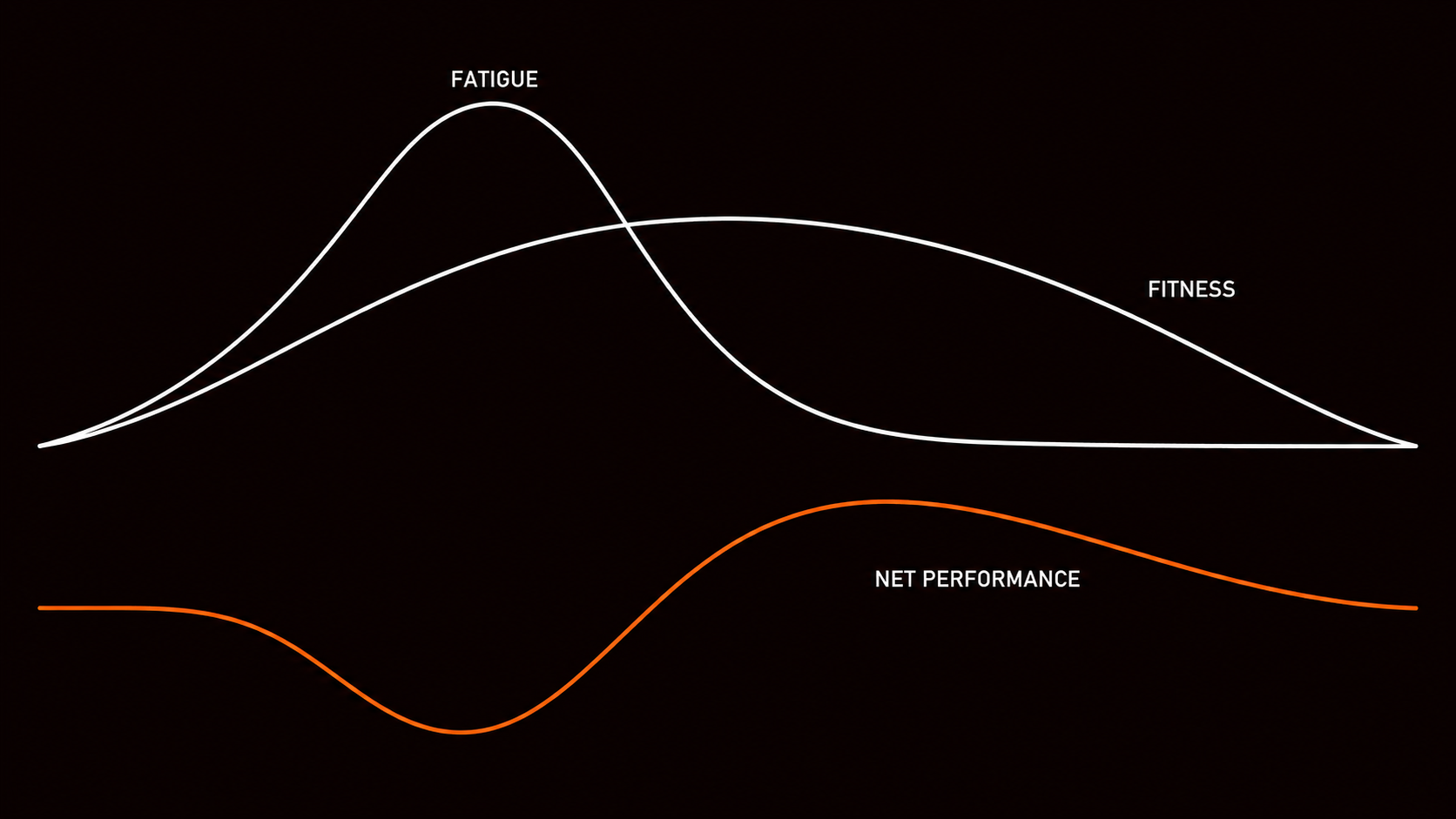
The fitness-fatigue model provides the structural framework for understanding why training does not produce linear improvement. (Morton et al. 1990) Every training stimulus produces two simultaneous biological responses: a positive fitness effect and a negative fatigue effect. Fitness, in this context, refers to the accumulation of physiological adaptations that increase capacity: greater mitochondrial density, improved metabolic flexibility, increased tendon and muscle resilience. Fatigue refers to the acute biological suppression of those same capacities: elevated stress hormones, glycogen depletion, disrupted autonomic balance, incomplete tissue repair.
These two components do not decay at the same rate. Fitness gains are slower to build and slower to dissipate. Fatigue accumulates quickly and, under adequate recovery, dissipates faster. This asymmetry is the structural basis of periodization: training phases generate both fitness and fatigue simultaneously, recovery phases allow fatigue to dissipate while fitness is largely preserved, and net performance peaks at the point where fitness minus fatigue is maximized.
THE FITNESS-FATIGUE MODEL
FITNESS COMPONENT
The positive biological adaptation from training: greater mitochondrial density, improved metabolic flexibility, increased structural resilience. Accumulates slowly. Decays slowly.
FATIGUE COMPONENT
The acute biological suppression produced by the same training: elevated stress hormones, glycogen depletion, disrupted autonomic balance, incomplete tissue repair. Accumulates quickly. Decays faster than fitness.
Durability, as defined here, is not a single peak of net positive performance. It is the sustained ability to maintain net positive performance across repeated training cycles, without the fatigue component progressively outpacing recovery. When that balance holds over months and training years, the athlete produces consistent, high-quality output. When it fails repeatedly because load exceeds recovery, net performance trends downward even as training volume stays constant or increases. Training harder for diminishing returns, effort that does not translate into improvement, output that feels inconsistent and unreliable: this is not a psychological problem. It is a specific failure mode in the fitness-fatigue balance, and it has identifiable causes.
Durability, as defined here, is not a single peak of net positive performance. It is the sustained ability to maintain net positive performance across repeated training cycles, without the fatigue component progressively outpacing recovery.
What Builds the Fitness Side of the Equation
Mitochondrial Density and the Aerobic Ceiling
The biological ceiling on sustainable aerobic output is set primarily by mitochondrial density: the number of mitochondria per unit of muscle tissue. (Holloszy 1967) Mitochondria are the cellular structures responsible for oxidative phosphorylation, the process by which the body produces the majority of its ATP through aerobic pathways. Higher mitochondrial density means more parallel ATP synthesis capacity, a higher absolute aerobic output, and a higher threshold at which anaerobic pathways must be recruited.
When work intensity exceeds the aerobic ceiling, the body recruits anaerobic glycolytic pathways to bridge the shortfall. This produces lactate accumulation, a steeper rise in perceived effort, and accelerated fatigue. From the perspective of the fitness-fatigue model, crossing the aerobic threshold repeatedly and prematurely increases the rate at which the fatigue component accumulates per session. An athlete with a higher aerobic ceiling can produce more absolute work at the same relative fatigue cost.
Mitochondrial density responds to a specific training stimulus: sustained aerobic work at or below the lactate threshold. This range, commonly referred to as zone 2 training, activates PGC-1alpha, a transcriptional coactivator that upregulates the expression of proteins involved in mitochondrial biogenesis. (Baar et al. 2002) (MacInnis & Gibala 2017) The adaptation is not rapid. Mitochondrial density increases over weeks and months of consistent aerobic training, and it declines with prolonged inactivity. Because it accumulates slowly and decays slowly, it sits squarely on the fitness side of the model.
01
Mitochondrial Density
Sets the aerobic ceiling. Built through sustained zone 2 aerobic training via PGC-1alpha activation. Accumulates slowly over weeks and months. Determines how much work can be produced before anaerobic pathways must compensate.
02
Metabolic Flexibility
Determines how efficiently the aerobic ceiling is used. The capacity to oxidize fat at moderate intensities, sparing glycogen. Built through aerobic training combined with appropriate dietary structure. Reduces glycogen cost per session and fatigue accumulation rate.
What Metabolic Flexibility Means for Sustainable Output
Mitochondrial density determines how high the aerobic ceiling is. Metabolic flexibility determines how efficiently that ceiling is used. Metabolic flexibility is the physiological capacity to shift primary fuel substrate between fat and carbohydrate in response to availability and demand. (Galgani et al. 2008) An athlete with high metabolic flexibility can sustain moderate-intensity work primarily on fat oxidation, keeping glycogen in reserve for high-intensity efforts that genuinely require it. An athlete with low metabolic flexibility recruits glycogen at intensities where fat oxidation should dominate, depleting stores faster and driving earlier fatigue.
The practical consequence of this difference is significant. (San-Millán & Brooks 2018) A metabolically flexible athlete trains at moderate intensity with a lower glycogen cost per session. He enters subsequent sessions with more complete fuel stores, produces work at equivalent absolute output with lower perceived effort, and accumulates fatigue more slowly across a training week. A metabolically inflexible athlete drains faster, recovers more slowly between sessions, and accumulates the fatigue component of the fitness-fatigue equation at a higher rate per unit of training load.
Metabolic flexibility is a product of chronic aerobic training combined with dietary patterns that do not chronically suppress fat oxidation pathways. It is not a fixed trait. It is a physiological state that responds to training and can be improved or degraded depending on how an athlete structures both work and nutrition.
An athlete with low metabolic flexibility recruits glycogen at intensities where fat oxidation should dominate, depleting stores faster and driving earlier fatigue.
Mitochondrial density responds to a specific training stimulus: sustained aerobic work at or below the lactate threshold.
What Drives the Fatigue Side of the Equation
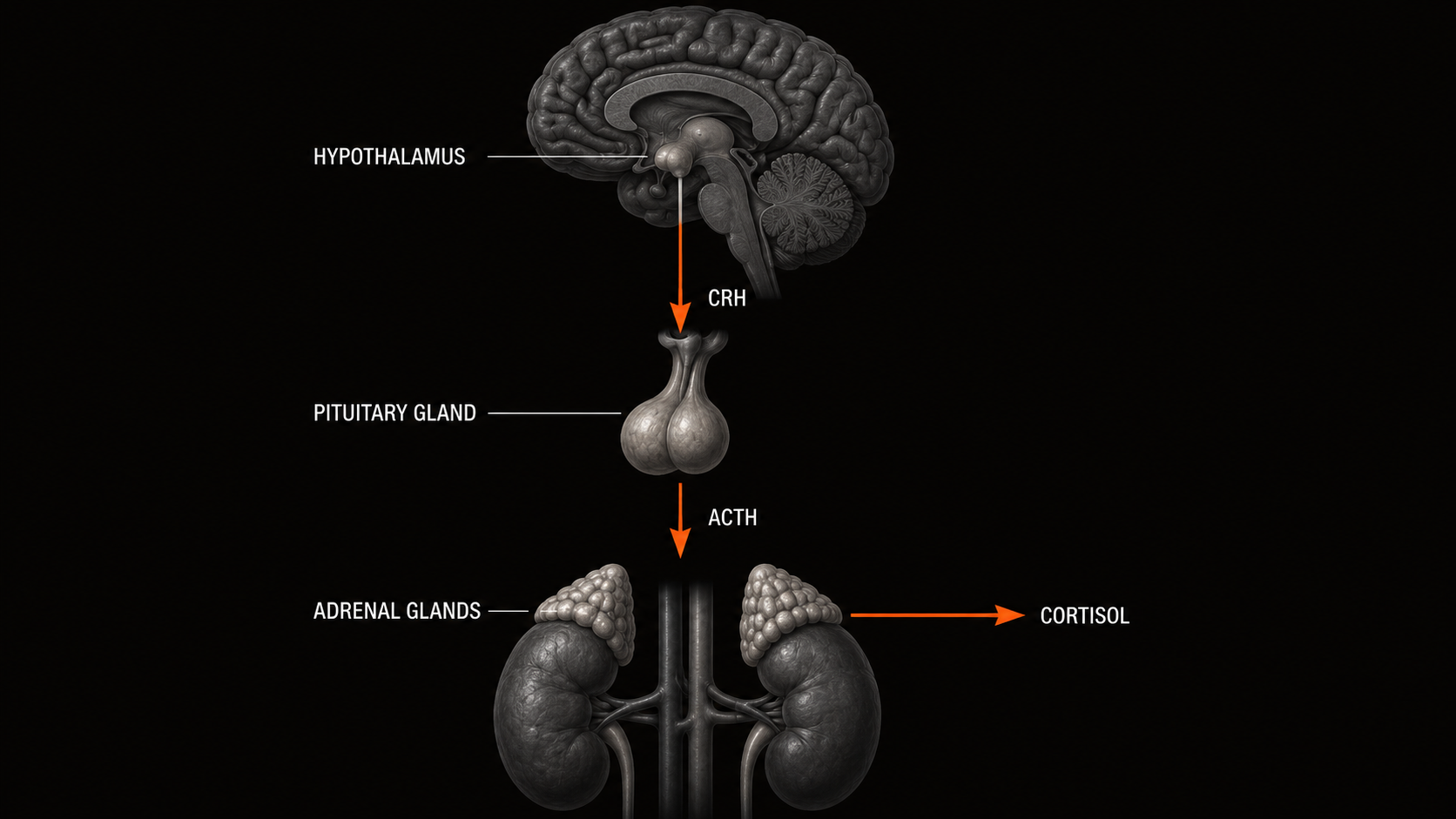
The HPA Axis and Cortisol
The hypothalamic-pituitary-adrenal axis is the body's primary stress-response system. (Meeusen et al. 2013) When a stressor is detected, the hypothalamus releases corticotropin-releasing hormone, which signals the pituitary to release adrenocorticotropic hormone, which in turn drives cortisol secretion from the adrenal cortex. Cortisol is an essential component of the acute stress response: it mobilizes fuel, reduces inflammation in the short term, and supports the physiological demands of effort.
Chronic cortisol elevation, which occurs when training load consistently exceeds recovery capacity, shifts the hormonal environment from adaptive to catabolic. Chronically elevated cortisol suppresses muscle protein synthesis, impairs glycogen resynthesis, degrades sleep architecture, and inhibits the anabolic signaling cascades that convert training stimulus into fitness adaptation. (Meeusen et al. 2013) In the context of the fitness-fatigue model, this means that chronic HPA dysregulation accelerates the fatigue component without proportionally increasing the fitness component. The inputs (training volume) remain constant; the biological return on those inputs declines because the recovery machinery is being systematically undermined.
Two distinct HPA dysregulation patterns appear in overtrained athletes. Sympathetic overtraining, more common in power and combat sports athletes, presents with sustained cortisol elevation, elevated resting heart rate, and disrupted sleep. Parasympathetic overtraining, more common in high-volume endurance athletes, presents with blunted cortisol response, chronically low resting heart rate, and persistent fatigue despite reduced training load. Both are regulatory failures, and both produce declining net performance, though through different mechanisms.
TWO HPA DYSREGULATION PATTERNS
SYMPATHETIC OVERTRAINING
More common in power and combat sports athletes. Presents with sustained cortisol elevation, elevated resting heart rate, disrupted sleep, and heightened sympathetic activity.
PARASYMPATHETIC OVERTRAINING
More common in high-volume endurance athletes. Presents with blunted cortisol response, chronically low resting heart rate, and persistent fatigue despite reduced training load.
Why Life Stress Counts as Training Stress
The HPA axis does not distinguish between sources of stress. A difficult week at work, a period of poor sleep, a family situation that requires sustained cognitive and emotional resources: each of these activates the same hormonal cascade as a hard training session. The relevant variable for recovery capacity is not training load in isolation. It is total allostatic load, the cumulative physiological cost across all stressors, regardless of their origin.
This matters significantly for the target reader. An athlete managing a demanding career alongside serious training has an HPA axis exposed to compounding inputs from multiple sources. Tracking training volume carefully while ignoring non-training stress produces a systematically incomplete picture of where recovery debt is accumulating. Athletes in this position often experience declining HRV during weeks with low training load and cannot explain why, because the explanation lies in the occupational and personal demands of those same weeks, not in the training log.
The practical implication is not to reduce training whenever life is demanding. It is to account for total load when interpreting the body's signals, to avoid compounding already elevated stress with training choices that further strain the HPA axis, and to recognize when recovery capacity is genuinely reduced rather than attributing all performance variation to motivation or mental readiness.
Chronic cortisol elevation, which occurs when training load consistently exceeds recovery capacity, shifts the hormonal environment from adaptive to catabolic.
The relevant variable for recovery capacity is not training load in isolation. It is total allostatic load, the cumulative physiological cost across all stressors, regardless of their origin.
Tracking the Balance: What the Data Actually Shows
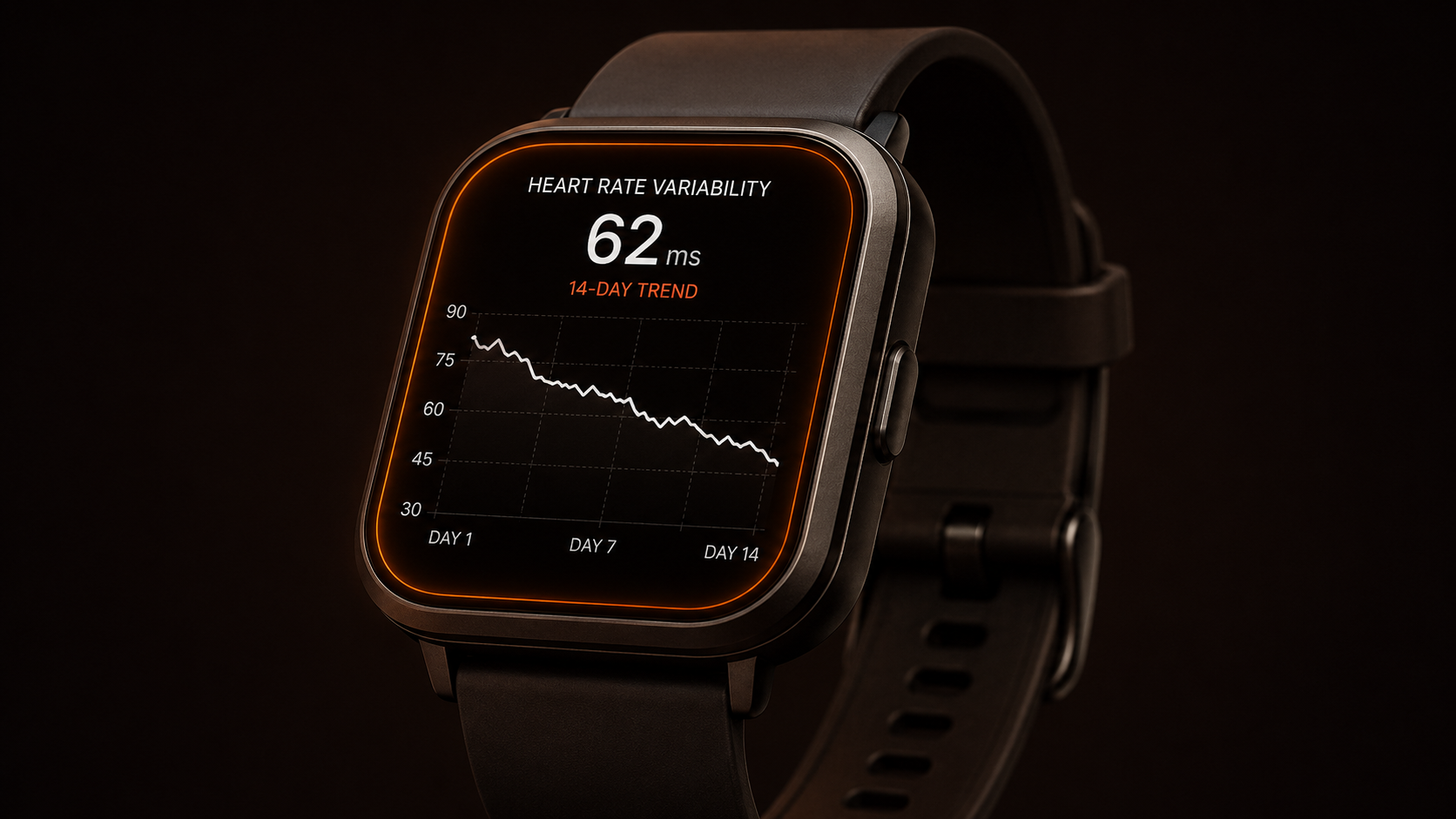
Heart Rate Variability as an Early Warning Signal in Athletes
Heart rate variability is the variation in time between successive heartbeats. It is measured in milliseconds and reflects the relative balance between sympathetic and parasympathetic autonomic activity. Higher HRV indicates greater parasympathetic (recovery) modulation; declining HRV indicates increasing sympathetic (stress) dominance. (Task Force 1996) At a population level, higher HRV is associated with greater aerobic fitness, better recovery capacity, and lower allostatic load.
A single HRV reading has limited predictive value. Day-to-day variation is normal and reflects acute changes in sleep quality, hydration status, and recent training. What is informative is trend: a sustained downward trajectory in HRV over seven to fourteen days, particularly when accompanied by a rising resting heart rate, indicates that the fatigue side of the fitness-fatigue balance is progressively outpacing recovery. (Plews et al. 2013) This pattern can precede subjective performance decline by days, providing a measurable early-warning signal for non-functional overreaching before it progresses to overtraining syndrome, which requires months to resolve.
HRV does not identify the cause of the imbalance. It indicates that an imbalance exists. Whether the driver is training load, sleep debt, life stress, or inadequate fueling requires a separate diagnostic process. The value of HRV monitoring is precisely this: it converts a subjective signal into a trackable biological pattern, which is more actionable than either dismissing the signal or attributing it to a motivation failure. For a fuller breakdown of the biological variables behind day-to-day readiness variation, see why some days feel effortless and others don't.
7–14
DAYS OF HRV DECLINE
Sustained downward HRV trend before subjective performance decline
24–48
HOURS MPS ELEVATED
Window during which muscle protein synthesis remains elevated post-training
2
HOURS PEAK RESYNTHESIS
Period of highest glycogen resynthesis rate following exercise
The Recovery Window: What Must Complete Between Sessions
Recovery is not a passive state between training sessions. It is a set of active biological processes that must run to completion for training adaptation to be captured. These processes begin during and immediately after training and extend for hours, with many completing during sleep. When a subsequent training session begins before these processes have completed, the net effect is less adaptation per unit of training stress, because the input exceeded the system's capacity to respond to it.
Muscle protein synthesis is the process by which damaged muscle proteins are replaced with new, stronger proteins in response to training stimulus. Following resistance training, synthesis rates increase significantly within hours and remain elevated for twenty-four to forty-eight hours, depending on training intensity and the individual's training status. (Phillips et al. 1997) Beginning a high-load session within this window, before synthesis has approached completion, imposes additional disruption on a repair process already underway and reduces net adaptation per unit of training stress.
Glycogen resynthesis rates are highest in the first two hours post-exercise, particularly when carbohydrate is available, and decline substantially in subsequent hours. (Ivy et al. 1988) Starting a subsequent session on partially depleted glycogen stores shifts the metabolic profile of that session: fat oxidation may be impaired by low glycogen availability, perceived effort rises at lower absolute output, and the training stimulus shifts toward what the depleted system can produce rather than what the adapted system would generate under adequate fueling.
HRV does not identify the cause of the imbalance. It indicates that an imbalance exists.
The athlete who recovers faster, in the functional sense, is not the one who feels ready sooner. It is the one whose biological recovery processes complete more efficiently per hour of recovery time, a consequence of aerobic fitness, adequate sleep architecture, and appropriate fueling, not of psychological willingness to return.
The athlete who recovers faster, in the functional sense, is not the one who feels ready sooner. It is the one whose biological recovery processes complete more efficiently per hour of recovery time.
Where Athletes Go Wrong With Durability
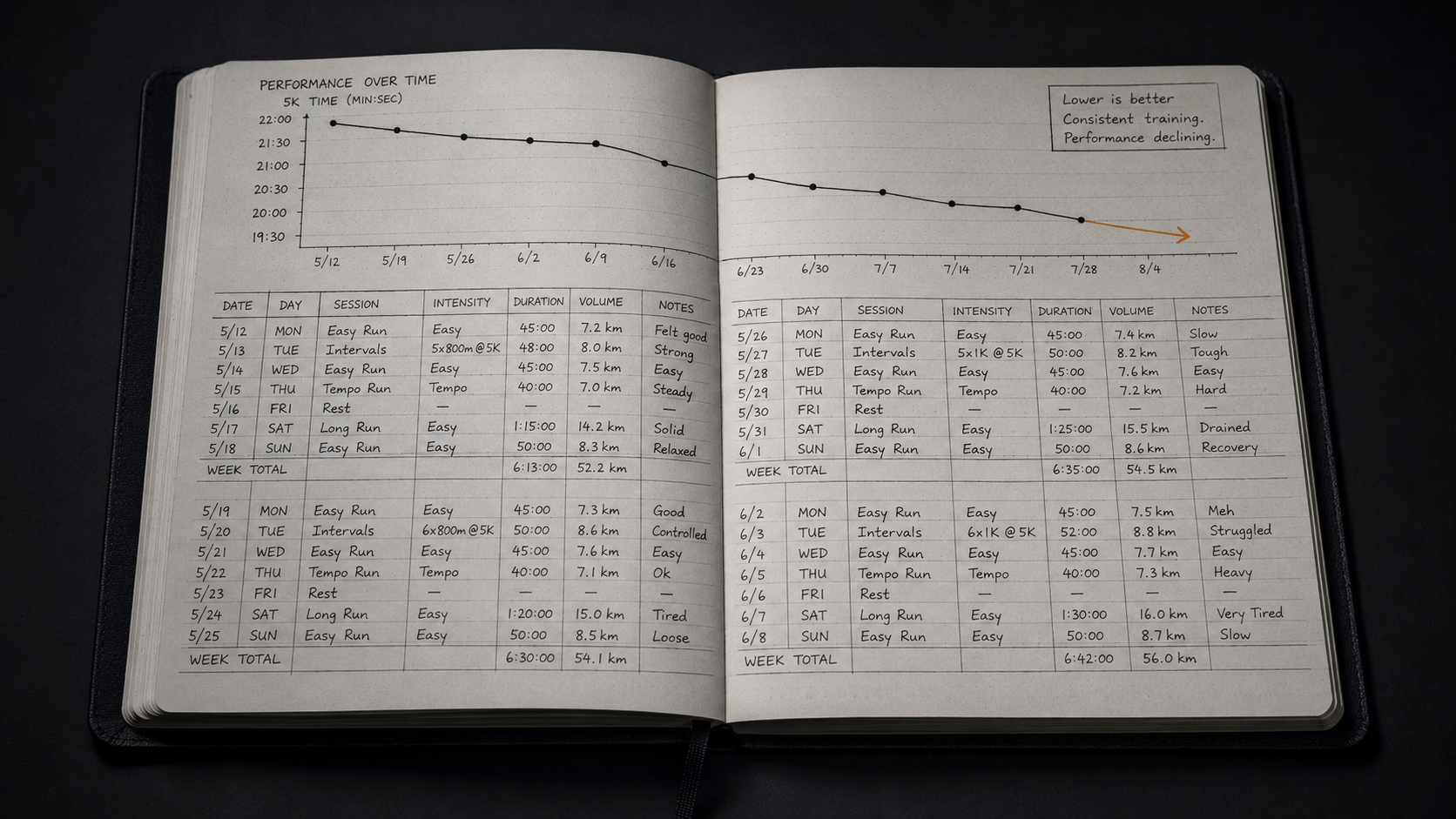
The most common failure mode in athletes pursuing durability is optimizing for peak output rather than consistent net-positive output. Training that pushes toward maximum intensity in every session generates a high fitness stimulus but also generates high fatigue. If fatigue does not adequately dissipate before the next session, net performance declines even as fitness continues to accrue. The athlete trains harder and experiences less performance improvement. The instinctive response, adding more effort or more sessions, further compounds fatigue accumulation without providing the recovery window required for the fitness component to express itself. This is a timing problem in the fitness-fatigue balance, not a deficit in effort or commitment.
The second common failure is treating training load as the only meaningful input to the recovery equation. Non-training stress, occupational demand, sleep shortfalls, and life load all activate the HPA axis and consume recovery capacity. An athlete who controls training volume carefully but carries significant non-training load is exposing his stress-response system to a total allostatic burden that his training log cannot account for. The result is a persistent disconnect between what the training log predicts and what the body actually produces. Understanding why daily readiness varies is the first step toward closing that gap.
The third failure is misinterpreting the phenomenology of fatigue accumulation as a motivational problem. When the fatigue component of the fitness-fatigue equation consistently outpaces recovery over multiple weeks, the subjective experience includes declining motivation, reduced drive, an aversion to the training that previously felt rewarding, and a sense that performance has become unreliable. This is not a character deficit. It is the predictable subjective expression of sustained autonomic and hormonal dysregulation: sympathetic dominance, elevated cortisol, and declining anabolic hormone levels. Pushing harder in this state further accelerates fatigue accumulation because the recovery infrastructure is compromised. The appropriate response is a recovery intervention targeting the HPA axis, sleep architecture, and total load, in that order. The clinical presentation of this failure mode is described in detail in the article on overtraining syndrome.
01
Optimizing for Peak Output
Training that maximizes intensity in every session generates high fatigue alongside fitness. Without adequate recovery, net performance declines even as fitness accrues.
02
Treating Training Load as the Only Input
Non-training stress activates the HPA axis and consumes recovery capacity. A training log that ignores life stress systematically underestimates total fatigue accumulation.
03
Misreading Fatigue as Motivation Failure
Declining drive and output under chronic fatigue accumulation are physiological outputs of autonomic and hormonal dysregulation, not character deficits. Pushing harder accelerates the condition.
This is a timing problem in the fitness-fatigue balance, not a deficit in effort or commitment.
What This Looks Like in Practice

Training Structure for the Aerobic Base
Zone 2 training, sustained aerobic effort below the lactate threshold maintained for forty-five to ninety minutes, is the primary structural input for building the mitochondrial density and fat oxidation capacity that expand the fitness side of the fitness-fatigue model. The adaptation it produces, greater mitochondrial density and improved fat oxidation capacity, is delivered at comparatively low fatigue cost. This means the fitness-fatigue ratio improves with each session: fitness accumulates, but the fatigue component generated per session is modest relative to the adaptation produced.
Athletes who train primarily at high intensity build fitness stimulus but also accumulate fatigue rapidly. Without an adequate aerobic base, high-intensity work extracts more from the recovery budget than it contributes to adaptation. Adding structured zone 2 training is not about doing easier sessions. It is about improving the biological return on total training investment by ensuring that the foundation on which high-intensity work sits is robust enough to support it. Three to four weekly sessions of sustained zone 2 work, over twelve to twenty-four weeks, produces the mitochondrial density and fat oxidation capacity that extends the aerobic ceiling and improves session-to-session fuel efficiency.
The athlete's practical task is not to replace high-intensity training with zone 2. It is to build the aerobic base to the point where high-intensity work is more efficiently supported, recovery between sessions is faster, and the fitness-fatigue balance can be maintained across longer training blocks without the fatigue component compounding.
45–90
MINUTES PER SESSION
Sustained aerobic effort below lactate threshold
3–4
SESSIONS PER WEEK
Weekly frequency for mitochondrial density adaptation
12–24
WEEKS TO ADAPT
Timeline for meaningful mitochondrial density and fat oxidation improvement
Substrate and Diet: How Fuel Choices Affect the Balance
Dietary composition affects the metabolic flexibility that determines fuel substrate at a given exercise intensity. Chronically high carbohydrate availability with low aerobic training produces a state in which the body's fat oxidation machinery is underutilized and undertrained. In this state, glycogen becomes the default fuel at intensities where fat oxidation would be more efficient, depleting stores faster and increasing the session-to-session recovery cost. The rate at which glycogen is depleted at different intensities is examined in detail in what actually happens when carbs run low.
The structural approach that supports metabolic flexibility is carbohydrate periodization: matching carbohydrate availability to training demand rather than maintaining constant high availability regardless of session type. The goal here is not carbohydrate restriction. Sessions that target aerobic adaptation benefit from lower pre-session carbohydrate availability, because the metabolic environment of low glycogen reinforces fat oxidation pathways and strengthens the fuel-switching capacity over time. Sessions that require high-intensity output depend on glycogen and should be adequately fueled.
Adding structured zone 2 training is not about doing easier sessions. It is about improving the biological return on total training investment by ensuring that the foundation on which high-intensity work sits is robust enough to support it.
The consequence of improved metabolic flexibility on the fitness-fatigue balance is direct: an athlete who burns less glycogen per session at equivalent moderate-intensity output preserves more for high-intensity efforts, enters subsequent sessions with more complete fuel stores, and reduces the fatigue cost of each session without reducing the fitness stimulus.
The consequence of improved metabolic flexibility on the fitness-fatigue balance is direct: an athlete who burns less glycogen per session at equivalent moderate-intensity output preserves more for high-intensity efforts, enters subsequent sessions with more complete fuel stores, and reduces the fatigue cost of each session without reducing the fitness stimulus.
Frequently Asked Questions
What is the fitness-fatigue model?
The fitness-fatigue model describes how every training session produces two simultaneous biological effects: a positive fitness component from adaptation and a negative fatigue component that temporarily suppresses performance. Fitness accumulates and dissipates slowly; fatigue accumulates and dissipates faster. Net performance at any point reflects the difference between the two.
Is durability the same as fitness?
No. Fitness describes the physiological adaptations built through training. Durability describes the ability to maintain net positive performance consistently over time. High fitness with chronically unmanaged fatigue produces inconsistent, unreliable output. Durability requires fitness but also depends on managing the fatigue component of the fitness-fatigue balance.
How does HRV reflect recovery status?
Heart rate variability reflects the balance between sympathetic and parasympathetic autonomic activity. Higher HRV indicates a recovered, parasympathetic-dominant state. A sustained downward trend over seven to fourteen days signals that fatigue is outpacing recovery in the fitness-fatigue balance. A single low reading is not reliably informative; trend over time is. For more on what drives daily HRV variation, see why some days feel effortless.
Why does life stress affect training recovery?
The HPA axis activates the same cortisol-releasing cascade in response to any stressor, regardless of source. Occupational demand, poor sleep, and psychosocial stress consume the same recovery capacity as training. When non-training stress is elevated, the recovery budget available for training adaptation is reduced even if training volume is unchanged.
What is metabolic flexibility and why does it matter for durability?
Metabolic flexibility is the capacity to shift primary fuel substrate between fat and carbohydrate in response to demand. A metabolically flexible athlete oxidizes predominantly fat at moderate intensities, sparing glycogen for high-intensity work. This reduces the fuel cost per session and the recovery burden between sessions, supporting the fitness-fatigue balance over time.
The Bottom Line
Durability is measurable. It is not a personality trait or a function of how much discomfort an athlete can tolerate. It is the biological condition that exists when the fitness component of the fitness-fatigue model consistently outpaces the fatigue component, session after session, week after week, training year after training year.
That condition is shaped by specific, trainable variables: mitochondrial density (built through consistent aerobic work), metabolic flexibility (built through training structure and appropriate carbohydrate periodization), HPA axis regulation (maintained by matching total load to recovery capacity), and sleep-dependent repair cycles (maintained by protecting the biological window in which adaptation completes). None of these are motivational variables. All of them respond to inputs that are under the athlete's control.
The most common way athletes erode durability without recognizing it is by treating each session as an isolated event to be maximized, rather than as one element in a sequence that must be managed for cumulative net output. The athlete who builds durability is building a system that compounds over time, not one that depletes in pursuit of peaks it cannot sustain.
The athlete who builds durability is building a system that compounds over time, not one that depletes in pursuit of peaks it cannot sustain.
Training builds both fitness and fatigue at the same time. Durability is what happens when recovery consistently keeps the balance positive.
The fitness side responds to consistent aerobic work, adequate fueling, and sleep-protected recovery cycles. The fatigue side responds to total load — not just training load. Managing both sides, not maximizing one, is what produces performance that compounds.
Mydos Performance
— Sustained Energy vs. Borrowed Energy Series —
The Fitness-Fatigue Model: What Durability Actually Means in Performance
Why repeatable output is the only standard that compounds over a training career
By Ricardo Londono · 2026