
The Energy System Model Most Athletes Learn Is Incomplete

When coaches first introduce the three energy systems explained in sports physiology, the standard presentation goes something like this: the body starts with phosphocreatine for explosive work, burns through those stores and shifts to glycolysis for sustained high-intensity effort, then eventually transitions to aerobic metabolism for prolonged exertion. This model is pedagogically convenient, creating a clear mental picture and mapping onto popular training frameworks. The problem is that it is mechanistically wrong.
The three energy systems do not operate in sequence. They operate in parallel. At rest, during a tempo run, and at the top of a maximal sprint, all three systems are contributing to ATP resynthesis simultaneously. Intensity and duration determine which system is contributing the most at any given moment, not which one has turned on. This distinction has consequences for how rest intervals are designed, why pacing feels the way it does, and what training stimuli are actually targeting. The cellular mechanism behind the most demanding of these efforts — the anaerobic energy system — is covered in depth in the adjacent article in this pillar.
Sequential Model (How Most Athletes Think)
Three phases. Each system waits its turn.
PCr fires first, then glycolysis takes over, then aerobic metabolism kicks in. One ends before the next begins.
Continuum Model (How It Actually Works)
Three systems. All active always. Proportion shifts with intensity.
All three systems run simultaneously at every intensity. Effort level determines which contributes most — not which one turns on.
This article works through the mechanism of each system, addresses the most common misapplication of energy system concepts, and builds toward the model that more accurately reflects the physiology: a continuum of overlapping contributions rather than a sequence of distinct phases. Four questions from readers searching this topic are worth noting upfront and will be answered directly in the sections that follow: what are the three main energy systems, what are the three different types of energy systems, what are the three energy systems and their basic functions, and what are the top three energy sources. The answers are mechanistic, not definitional.
The three energy systems do not operate in sequence. They operate in parallel.
ATP: The Only Currency the Muscle Accepts
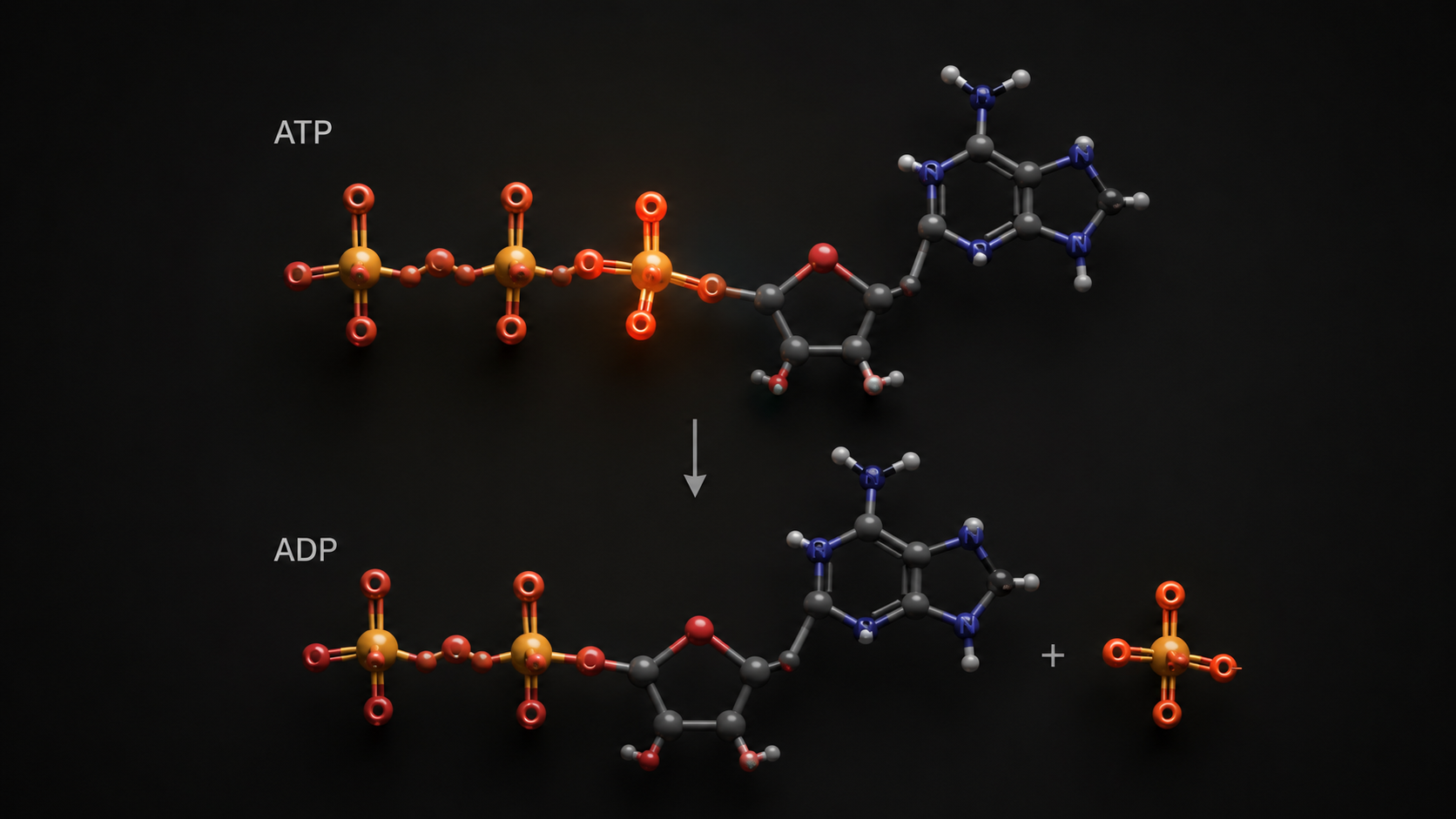
Every contraction, regardless of intensity or duration, requires adenosine triphosphate. ATP is a small molecule that releases energy when its terminal phosphate bond is broken, converting ATP to ADP (adenosine diphosphate) and a free phosphate group. That released energy powers the molecular machinery of muscle contraction. No other molecule performs this function in the working muscle.
The central constraint is that muscle tissue stores a very small amount of ATP at any given time, enough to sustain roughly two to three seconds of maximal effort before stores are exhausted. This is not a design flaw; it is a consequence of ATP's molecular weight and the energetic cost of maintaining large stores. The implication is that continuous ATP resynthesis is non-negotiable. All three energy systems exist to solve the same problem: rebuilding ADP back into ATP fast enough to sustain contraction.
THE THREE SYSTEMS ON ONE AXIS
Phosphocreatine System
Highest rate. Lowest capacity. Stores depleted in 6–10 seconds at maximal effort.
Glycolytic System
Intermediate rate. Intermediate capacity. Dominant for 30 seconds to 2 minutes.
Aerobic System
Lowest rate. Highest capacity. Sustains output for hours.
Mitochondria play a central role in this resynthesis process under aerobic conditions. They are the primary site of oxidative phosphorylation, the mechanism by which oxygen is used to produce the majority of the body's ATP during sustained exercise. The role of mitochondria in ATP production during aerobic work, and how endurance training increases mitochondrial density, is addressed in detail in the section on the aerobic system. Before that, the two systems that do not require oxygen deserve their own treatment, because they operate through fundamentally different mechanisms and on fundamentally different timescales.
The three systems can be understood along a single axis: rate versus capacity. Rate refers to how quickly a system can regenerate ATP per unit of time. Capacity refers to how long the system can sustain that rate before substrate depletion or accumulating byproducts limit further output. The phosphocreatine system has the highest rate and the lowest capacity. The aerobic system has the lowest rate and the highest capacity. The glycolytic system sits between them on both dimensions.
All three energy systems exist to solve the same problem: rebuilding ADP back into ATP fast enough to sustain contraction.
System 1: What Is Phosphocreatine and How Does It Work

Phosphocreatine (PCr), often abbreviated as PCr or creatine phosphate, is a compound stored directly in muscle tissue alongside ATP. Its structure includes a high-energy phosphate bond that can be rapidly donated to ADP to regenerate ATP. The reaction is catalyzed by an enzyme called creatine kinase and proceeds in milliseconds, without any requirement for oxygen, glucose, or multi-step enzymatic pathways.
This makes the phosphocreatine system the fastest ATP-producing mechanism available to the muscle. From the onset of a maximal effort, PCr is providing ATP before the glycolytic and aerobic systems have had time to accelerate. The tradeoff is that PCr stores in a given muscle are finite and small. Under maximal intensity conditions, PCr is substantially depleted within six to ten seconds. This is not a failure of training; it is the nature of the substrate. It explains why truly maximal efforts, whether a 40-meter sprint, a maximal clean, or an explosive punch combination, degrade in quality after the first several seconds even in well-trained athletes.
6–10 SEC
Maximum duration at full power output
~70%
PCr restored after 30 seconds of rest
3–5 MIN
Time to full PCr restoration
Source: Bogdanis et al. 1996
PCr stores do replenish during rest and recovery. Research examining PCr resynthesis kinetics following maximal sprint exercise indicates that stores recover to approximately 70 percent of baseline within 30 seconds of reduced-intensity activity and approach full restoration within three to five minutes (Bogdanis et al. 1996). This recovery is largely aerobic: the oxidative system drives the reconversion of creatine back to phosphocreatine. The dependence of PCr recovery on aerobic metabolism is one reason an aerobic base supports power-based performance.
PCr stores are substantially depleted within six to ten seconds.
Understanding what is phosphocreatine at a mechanistic level is also important for interpreting creatine supplementation, which increases resting PCr stores in muscle tissue. That is a topic addressed in a companion article; for this discussion, the mechanism of PCr resynthesis during exercise is the relevant layer.
The phosphocreatine system is not the only anaerobic system. When effort duration extends beyond the window PCr can support, glycolysis takes over as the primary fast ATP-producing pathway.
The dependence of PCr recovery on aerobic metabolism is one reason an aerobic base supports power-based performance.
System 2: How Glycolysis Produces ATP

Glycolysis is a ten-step enzymatic process that breaks one molecule of glucose down into two molecules of pyruvate, generating a net yield of two ATP molecules in the process. When glycogen stored in muscle tissue is the substrate rather than circulating blood glucose, the yield increases to three ATP. The process occurs in the cytoplasm of the cell, requires no oxygen, and can sustain ATP production at a rate substantially higher than the aerobic system can maintain under similar time constraints.
Understanding how glycolysis produces ATP requires distinguishing the inputs from the outputs. Glucose or glycogen enters the pathway. The energy released during the stepwise breakdown of this substrate is captured as ATP and as NADH, an electron carrier. At moderate to high intensities, when oxygen delivery is insufficient to fully process the NADH produced, pyruvate accepts the electrons and is reduced to lactate. This step is not a system failure; it is an important mechanism for regenerating the electron carrier so glycolysis can continue producing ATP.
4.1
Input
Glucose (from blood) or glycogen (from muscle stores). One molecule of glucose enters the ten-step pathway.
4.2
Process
Ten enzymatic steps in the cytoplasm. No oxygen required at any step. Proceeds rapidly under high-intensity conditions.
4.3
Output
2 ATP from glucose; 3 ATP from glycogen. Plus NADH electron carriers for further downstream processing.
4.4
Byproduct at High Intensity
Pyruvate converts to lactate when oxygen delivery is insufficient, regenerating NADH so glycolysis can continue producing ATP without stopping.
The glycolytic system becomes the dominant ATP-producing pathway during high-intensity efforts lasting roughly 30 seconds to two minutes. Within this window, the combination of high ATP production rate and glycogen availability allows substantial work output. The system degrades not because glycolysis itself stops, but because the environment it creates, specifically the accumulation of hydrogen ions from concurrent ATP hydrolysis, progressively impairs the enzymatic machinery that drives contraction. A full account of what happens to effort quality across this window — and what the cellular experience of running into glycolytic failure actually involves — is in the adjacent article in this pillar.
The role of hydrogen ions in limiting glycolytic performance connects directly to one of the most widespread misconceptions in exercise physiology: the idea that lactic acid is responsible for the burning sensation in working muscles.
This step is not a system failure; it is an important mechanism for regenerating the electron carrier so glycolysis can continue producing ATP.
Lactate Is Not Lactic Acid, and It Is Not Waste

Lactic acid and lactate are not the same compound, and the distinction is not semantic. Lactic acid (C3H6O3) has a pKa of approximately 3.86, meaning it dissociates almost completely into lactate and a hydrogen ion at the physiological pH of working muscle tissue, which is maintained near 7.0. At this pH, lactic acid is present in negligible concentrations. What glycolysis produces in practice is lactate, not lactic acid (Robergs et al. 2004).
The burning sensation and impaired contractile function experienced during intense exercise are associated with rising intracellular acidity, specifically the accumulation of hydrogen ions. These hydrogen ions derive primarily from ATP hydrolysis, which occurs throughout the muscle whenever ATP is consumed, and from the dissociation of carbonic acid during CO2 clearance. The hydrogen ions that impair enzyme function are produced whether or not lactate is present. Lactate does not generate the H+ that drives acidosis; it is produced alongside the conditions that do (Robergs et al. 2004).
The Popular Model
Lactic acid = the burn, a waste product
Lactic acid builds up during intense exercise, causing the burning sensation and accumulating in the muscle as waste.
The Actual Mechanism
Lactate = fuel. H+ ions cause acidosis.
Lactate is transported to oxidative fibers, cardiac muscle, and the liver for energy. H+ from ATP hydrolysis drives the acidic environment.
Lactate's actual metabolic role is that of a fuel and a signaling molecule. It is not stored as waste. The intracellular lactate shuttle describes how lactate produced in fast-twitch, glycolytic fibers is transported across cell membranes and taken up by adjacent oxidative fibers, cardiac muscle, and the liver (Brooks 2000). In these tissues, lactate is converted back to pyruvate and enters the Krebs cycle as a carbon substrate for aerobic ATP production. Training improves both the rate at which lactate is produced and the rate at which it is cleared and oxidized. The practical consequence is that a better-trained athlete maintains higher absolute workloads before the hydrogen ion environment becomes contractile-function limiting, not because they produce less lactate, but because their lactate clearance and oxidation capacity is higher (Brooks 2018).
The burning sensation and impaired contractile function experienced during intense exercise are associated with rising intracellular acidity, specifically the accumulation of hydrogen ions.
The misidentification of lactate as the cause of fatigue has persisted in part because lactate and acidosis co-occur during intense exercise. The correlation is real, but the mechanism is not what the popular model assumes. How the H+ environment interacts with central nervous system fatigue signaling is an additional layer of the fatigue mechanism that compounds the peripheral effects described here.
Training improves both the rate at which lactate is produced and the rate at which it is cleared and oxidized.
System 3: How the Aerobic Energy System Works

The aerobic system, also called the oxidative system, produces ATP through a process that requires oxygen as the final electron acceptor. This occurs inside the mitochondria, the organelles distributed throughout skeletal muscle fibers in variable density depending on fiber type and training history. The aerobic system can oxidize carbohydrates, fats, and to a limited extent protein, which gives it a broad substrate base and a theoretically large energy capacity.
Understanding how the aerobic energy system works requires following the substrate through its pathway. Glucose begins glycolysis in the cytoplasm, generating pyruvate. Under aerobic conditions, pyruvate is transported into the mitochondria and converted to acetyl-CoA. Acetyl-CoA enters the Krebs cycle, a series of reactions that extract electrons and deposit them onto NADH and FADH2, the electron carrier molecules. These carriers then donate their electrons to the electron transport chain, a series of protein complexes embedded in the inner mitochondrial membrane. As electrons move through the chain, protons are pumped across the membrane, creating a gradient that drives ATP synthesis through ATP synthase. The specific role oxygen plays in this terminal step — accepting electrons to form water — is what gives oxidative phosphorylation both its name and its rate limit.
THREE AEROBIC FUEL PATHWAYS
Carbohydrates
Glucose → Pyruvate (glycolysis) → Acetyl-CoA (mitochondria) → Krebs cycle → Electron transport chain. Faster delivery than fat.
Fats
Fatty acids → Beta-oxidation → Acetyl-CoA → Krebs cycle → Electron transport chain. Higher ATP yield per gram; slower to mobilize. The shift in substrate utilization across intensities is covered in detail in energy in fat vs. carbohydrate.
Common Endpoint
Both substrates feed acetyl-CoA into the same downstream machinery. Fat yields more ATP per gram; carbohydrate delivers it faster. Intensity determines the proportion of each.
The total ATP yield from one glucose molecule through complete aerobic oxidation is approximately 30 ATP, compared to the two or three produced by glycolysis alone, a substantially higher output. Fatty acid oxidation follows a different pathway, beta-oxidation, which converts fatty acids into acetyl-CoA units before Krebs cycle entry. Fat yields more ATP per gram than carbohydrate but requires more oxygen per unit of ATP produced and cannot sustain ATP delivery as rapidly.
The aerobic system has the lowest rate of ATP production but the highest capacity. It can sustain output for hours, provided substrate availability and oxygen delivery are adequate. Its primary limitation during high-intensity exercise is not oxygen delivery per se, but the rate at which it can regenerate ATP relative to the demand created by fast-twitch fiber recruitment. When demand exceeds aerobic supply at high intensities, the glycolytic system accelerates to bridge the gap.
More mitochondria per fiber means a greater proportion of any given workload can be met aerobically, reducing the relative glycolytic contribution and the associated accumulation of hydrogen ions.
Endurance training increases mitochondrial density in skeletal muscle, expanding the aerobic system's capacity to produce ATP at any given workload (Holloszy and Coyle 1984). More mitochondria per fiber means a greater proportion of any given workload can be met aerobically, reducing the relative glycolytic contribution and the associated accumulation of hydrogen ions. This is the primary physiological adaptation underlying improved endurance performance with training, and it is one reason that aerobic base development is mechanistically relevant for athletes primarily focused on power and combat sports. The specific training stimuli that drive mitochondrial biogenesis — and how to structure them without compromising power adaptation — are covered in how to increase mitochondrial density.
The Energy Continuum: How Energy Systems and Exercise Interact

The energy continuum model states that all three energy systems contribute to ATP resynthesis simultaneously during all forms of exercise and rest. This is not a matter of interpretation; it is a consequence of how each system operates. The aerobic system does not wait for the phosphocreatine and glycolytic systems to finish before it turns on. It runs continuously, even during maximal sprint efforts, though its rate of ATP contribution cannot meet the full demand of maximum-intensity work.
The evidence for this comes from measurement of energy system interaction and relative contribution during maximal exercise across different durations. Research in this area consistently shows that even during very short maximal efforts, the aerobic system is providing a portion of the ATP being used, and that during prolonged submaximal exercise, the PCr and glycolytic systems continue to contribute alongside the aerobic pathway (Gastin 2001). The proportion shifts continuously rather than discretely.
7.1
At Rest
Aerobic system provides essentially all ATP. PCr and glycolytic contributions negligible. All three systems are active.
7.2
Low Intensity
Aerobic dominant. Glycolytic minor contributor. PCr maintained near resting levels. Fat oxidation provides majority of substrate.
7.3
High Intensity
Glycolytic contribution rises sharply. Aerobic still fully active. PCr contributes significantly during high-intensity bursts.
7.4
Maximal Effort (<10 sec)
PCr dominant. Glycolysis accelerating rapidly. Aerobic contributing but unable to meet full demand alone. All three systems running.
The practical implication of this continuum model for understanding how energy systems and exercise relate is that no training session, regardless of intensity, is purely anaerobic or purely aerobic. A maximal effort involves all three systems; the aerobic system contributes less to that effort in proportion but is never absent. A long, low-intensity effort involves all three systems; the PCr and glycolytic systems contribute less in proportion but are never switched off. This means training language like "aerobic training" or "anaerobic training" is best understood as describing which system is being stressed toward its limit, not which systems are active. The degree to which an athlete's body can shift fluidly between substrates at different intensities is what determines metabolic flexibility — a distinct but related concept covered separately.
The proportion shifts continuously rather than discretely.
There is also the temporal dimension. At the onset of any effort, including a long run, PCr provides the immediate ATP while the aerobic system accelerates to meet demand. This lag period, the oxygen deficit, represents the time between when oxygen consumption rises and when it reaches a steady state sufficient to meet the effort's full demand. During this window, the glycolytic system contributes substantially, even during efforts that are sustained aerobically for the following hour. Every exercise session, regardless of its dominant character, begins with an anaerobic contribution that the continuum model accounts for and the sequential model does not.
Every exercise session, regardless of its dominant character, begins with an anaerobic contribution that the continuum model accounts for and the sequential model does not.
How Intensity Determines Which System Dominates, and What Burns

Intensity determines the proportion of each system's contribution to ATP supply, and it simultaneously determines which substrates are being oxidized. These two variables move together because the systems differ in both their rate and their substrate preferences. At low intensities, roughly 50 percent of maximal oxygen uptake and below, the aerobic system supplies the vast majority of ATP. Fat oxidation accounts for a large proportion of the substrate being used. The glycolytic system is active but contributes minimally, and PCr is largely maintained at resting levels. As intensity rises, carbohydrate oxidation increases proportionally while fat oxidation decreases. Research on substrate utilization during exercise identifies a crossover point, the intensity at which energy from carbohydrate oxidation surpasses energy from fat oxidation. This crossover typically occurs in the moderate-intensity range, somewhere between 50 and 70 percent of VO2max in trained individuals, though the precise point varies with training status and dietary history (Brooks and Mercier 1994).
INTENSITY AND FUEL SELECTION
Low Intensity (<50% VO2max)
Aerobic system dominant. Fat oxidation provides majority of substrate. PCr largely maintained at resting levels.
Moderate Intensity — Crossover Zone (~50–70% VO2max)
Carbohydrate oxidation overtakes fat oxidation. Glycolytic contribution rises. Aerobic system still dominant.
High Intensity (>70% VO2max)
Glycolytic system dominant. Rapid H+ accumulation begins limiting sustainable workload. Fat oxidation continues at smaller proportion.
Maximal Effort (<10 sec)
PCr dominant. All three systems active. Substrate is primarily phosphocreatine and muscle glycogen. Glycolysis accelerating rapidly.
Above the crossover point, the glycolytic system's contribution to ATP supply grows substantially. Hydrogen ion accumulation begins to limit sustainable workload. PCr contribution also increases during bursts within this range. At truly maximal intensities lasting under 10 seconds, PCr is providing the dominant ATP contribution, glycolysis is accelerating rapidly, and the aerobic system is contributing a smaller proportion than at any other intensity. None of these proportions are binary; none involve the other systems switching off.
The practical takeaway from the continuum model applied to intensity is this: the metabolic consequences of effort are predictable from the underlying physiology. The burning sensation in high-intensity work follows from H+ accumulation, not from any intrinsic property of working hard. The relatively easy feeling of low-intensity effort follows from the aerobic system's high efficiency at those workloads, not from the body choosing a different mode. Training at different intensities produces different adaptations because it pushes different systems toward their limits.
The metabolic consequences of effort are predictable from the underlying physiology.
Where Energy System Thinking Goes Wrong

The sequential model produces predictable errors when athletes and coaches apply it to training design and interpretation of effort. These errors follow directly from the flawed model. Recognizing them is also how overtraining patterns related to energy system mismanagement develop.
Error 1
Training Energy Systems in Isolation
Neglecting aerobic base reduces PCr recovery between power bouts. The aerobic system is always relevant to power athletes — it drives PCr replenishment during rest intervals.
Error 2
Blaming Lactate for Soreness
DOMS peaks 24–72 hours post-exercise. Lactate clears within approximately one hour. The two are not causally connected. DOMS is caused by mechanical damage, not acid accumulation.
Error 3
Binary Substrate Thinking
No intensity exclusively burns fat or carbohydrate. The crossover is a proportion shift, not a binary switch. Fat oxidation continues even during high-intensity work.
Error 4
Overestimating PCr Duration
The 6–10 second window is real. Explosive quality degrades after this regardless of fitness level. Pacing errors often follow from expecting maximal power output to persist beyond this window.
The first error is treating energy system training as exclusive. Athletes focused on glycolytic conditioning sometimes neglect aerobic base development under the assumption that they are training a different system. The aerobic system is not being bypassed during high-intensity work; it is contributing to ATP supply and is the primary driver of PCr recovery between bouts. An aerobic base improves the rate at which PCr is restored during rest intervals, which directly supports the quality of subsequent maximal efforts. Neglecting aerobic development does not spare the aerobic system from being relevant to power and speed training.
The second error is attributing delayed onset muscle soreness to lactate accumulation. Delayed onset muscle soreness, which peaks 24 to 72 hours after exercise, is primarily caused by mechanical damage to muscle fibers and the subsequent inflammatory response. Lactate returns to baseline within approximately one hour of exercise cessation in most circumstances (Gladden 2004), well before the delayed soreness appears. The persistence of soreness for days after lactate is gone reveals that the two are not causally connected.
Neglecting aerobic development does not spare the aerobic system from being relevant to power and speed training.
The third error is assuming that low-intensity training exclusively burns fat while high-intensity training exclusively burns carbohydrates. Fat is the dominant substrate at low intensities, but carbohydrate oxidation is always occurring. The crossover is a proportion shift, not a binary switch. Similarly, high-intensity training does not burn exclusively carbohydrate; it shifts the proportion substantially toward carbohydrate while fat oxidation continues at an absolute level. How this proportion shift affects energy availability across a training session — including the downstream effects on performance when glycogen availability falls — is covered in the aerobic conditioning and oxygen delivery article.
The fourth error is overestimating the duration of phosphocreatine-dominant output. The six-to-ten second window is real. Athletes who believe their explosive quality persists longer than this often pace incorrectly in competition, starting efforts at genuinely maximal intensity and expecting to maintain quality well past the PCr window. The degradation that follows is a predictable consequence of PCr depletion.
Applying the Continuum Model

The continuum model provides a principled basis for reasoning about training and effort without prescribing specific protocols. Given how the three systems work, certain choices follow from the mechanism rather than from convention. Rest interval length in power-based training can be reasoned from the PCr replenishment timeline. If full PCr restoration requires approximately three to five minutes, then rest intervals substantially shorter than this will reduce PCr availability for subsequent maximal-intensity efforts. Whether shorter rest intervals are appropriate depends on what quality is being trained: if the goal is maximal power output, longer rest supports that goal; if the goal is adaptation to repeated sprint recovery, shorter rest creates the relevant stress. The physiology does not specify which goal to pursue; it specifies the consequence of each interval length. The fitness-fatigue model provides the framework for structuring these intervals across a training block without accumulating residual fatigue.
Pacing awareness follows from understanding the oxygen deficit and the PCr window. The degradation of effort quality at the start of the third round of a hard sparring session, or halfway through a repeated sprint protocol, is predictable from the cumulative depletion of PCr and glycogen and the accumulating H+ environment. This does not mean the decline is fixed or untreatable; it means the mechanism is understood, which is a prerequisite for addressing it.
Aerobic base investment is mechanistically warranted for athletes whose primary training is in combat sports or strength and power. The aerobic system drives PCr replenishment, supports lactate clearance and oxidation, and can be developed through training that does not interfere with power adaptation when volume and intensity are managed appropriately. The case for aerobic conditioning in power athletes is derived directly from the role the aerobic system plays in between-bout recovery.
The aerobic system drives PCr replenishment, supports lactate clearance and oxidation, and can be developed through training that does not interfere with power adaptation when volume and intensity are managed appropriately.
Frequently Asked Questions
What are the three main energy systems?
The three main energy systems are the phosphocreatine system, the glycolytic system, and the aerobic (oxidative) system. Phosphocreatine supports maximal efforts up to ten seconds. Glycolysis produces ATP from glucose without oxygen, sustaining efforts of 30 seconds to two minutes. The aerobic system uses oxygen to produce ATP from carbohydrate and fat, supporting sustained exercise.
Do the three energy systems work one at a time or simultaneously?
All three energy systems operate simultaneously at all times, including during rest. What changes during exercise is the proportion each system contributes to total ATP production. At high intensities, the phosphocreatine and glycolytic systems contribute proportionally more. At low intensities, the aerobic system supplies the vast majority of ATP. The systems do not switch on and off.
Is lactate the same as lactic acid?
Lactate and lactic acid are chemically distinct compounds. Lactic acid dissociates almost completely into lactate and a hydrogen ion at the pH of working muscle, meaning free lactic acid is present in negligible concentrations during exercise. The burning sensation in working muscle is caused by hydrogen ion accumulation and the resulting drop in intracellular pH, not by lactate itself.
What are the three energy systems and their basic functions?
The phosphocreatine system regenerates ATP instantly, supporting up to ten seconds of maximal effort. Glycolysis breaks glucose or glycogen into ATP without oxygen, sustaining high-intensity work for up to two minutes. The aerobic system uses oxygen to oxidize carbohydrate and fat, producing the highest ATP yield per substrate and supporting sustained exercise from minutes to hours.
What is substrate utilization during exercise?
Substrate utilization during exercise refers to which fuel sources the body draws on to produce ATP. At lower intensities, fat oxidation provides the majority of energy through the aerobic system. As intensity rises, carbohydrate oxidation increases proportionally. The crossover point is the exercise intensity at which energy from carbohydrate exceeds energy from fat, typically occurring in the moderate-intensity zone.
The Bottom Line
The sequential energy system model is a teaching tool that became a default framework, and applying it too literally produces predictable misunderstandings about effort, fatigue, and training design. The more accurate model is a continuum: all three systems are always operating, with intensity and duration continuously adjusting the proportion each contributes to ATP supply.
What this model provides is a basis for reasoning rather than a prescription. When an effort degrades after ten seconds of maximal output, the mechanism is PCr depletion. When the burning sensation appears in the third minute of a hard round, the mechanism is rising H+ concentration from rapid glycolytic activity and ATP hydrolysis. When fat is the primary substrate at low intensity, it is because the aerobic system's rate of ATP production is sufficient at that workload and fat oxidation carries the carbon load efficiently.
Each of those statements is predictable from the physiology. The value of understanding how the three energy systems work together is not in memorizing pathways; it is in having a model that generates accurate predictions about what happens to effort under different conditions.
Your body runs all three energy systems simultaneously. Intensity and duration determine which one contributes most.
The sequential model makes energy physiology easy to teach. The continuum model makes it possible to apply. Understanding the difference changes how rest, pacing, and training structure are reasoned about.
Mydos Performance
— Cellular Energy & Fuel Systems Series —
The Three Energy Systems Explained
How the body produces and deploys ATP across three overlapping pathways
· By Ricardo Londono, MD/PhD ·