
The Metric Most Athletes Train Around Misses the Real Limit

Aerobic capacity, as most athletes understand it, is a cardiovascular number. It describes how much oxygen the heart and lungs can deliver to working muscle per minute, and the standard metric used to track it, VO2max, reflects this delivery ceiling. This framework is accurate as far as it goes, but it describes only one side of the system that determines how much work a person can sustain.
Mitochondrial density in skeletal muscle is the other side. It describes not how much oxygen arrives at the muscle, but what the muscle can do with that oxygen once it gets there. Two athletes with identical VO2max values can have meaningfully different sustainable power outputs if one has substantially higher mitochondrial density in the working muscle fibers. The binding constraint on sustained effort is not always in the cardiovascular system. It can sit downstream, inside the muscle cells themselves.
Most training programs are built around the cardiovascular model. Volume, zones, heart rate targets, and VO2max tests all point to the delivery side of the equation. Understanding what the utilization side is, how it works, and what changes it requires starts at the cell level.
Two athletes with identical VO2max values can have meaningfully different sustainable power outputs if one has substantially higher mitochondrial density in the working muscle fibers.
What Mitochondria Do and Why Density Is the Variable That Matters

Mitochondria in Muscle Cells: The Site of Aerobic Work
Mitochondria are organelles found in nearly every cell in the body, but in skeletal muscle they serve a specific and central role: they are where aerobic ATP production occurs. Through oxidative phosphorylation, mitochondria use oxygen to convert substrates, primarily carbohydrates and fats, into adenosine triphosphate, the molecule that powers muscle contraction. The pathways that do not require oxygen (creatine phosphate and glycolysis) produce ATP faster, but their capacity is limited and cannot sustain effort beyond short durations. Anything lasting longer than a few seconds depends substantially on oxidative phosphorylation (Holloszy 1967).
The density of mitochondria per unit volume of muscle fiber sets how much aerobic ATP production the fiber can support at any given moment. A fiber with higher mitochondrial density can sustain a higher absolute work rate without recruiting anaerobic pathways. A fiber with lower density reaches its aerobic ceiling at a lower work rate and begins relying more heavily on glycolysis to fill the gap. This is why mitochondrial density functions as a performance variable independent of cardiovascular fitness: it sets the oxidative ceiling within the muscle itself.
Type I (slow-twitch) muscle fibers have inherently higher mitochondrial density than Type II fibers, which is the primary structural reason they are more fatigue-resistant during sustained effort. Type II fibers adapt their mitochondrial density substantially with appropriate aerobic training, narrowing this gap. Across individuals, trained athletes consistently show higher mitochondrial density than untrained people of comparable cardiovascular fitness, and this structural difference is one of the strongest predictors of endurance capacity (Holloszy and Coyle 1984).
TYPE I FIBER
High Mitochondrial Density
Preferentially oxidizes fat, highly fatigue-resistant during sustained effort. The default fiber type for aerobic work.
TYPE II FIBER
Lower Mitochondrial Density by Default
Relies more heavily on glycolysis, fatigues faster at sustained effort. Adapts substantially with consistent aerobic training.
Mitochondrial Efficiency: How Much Work Your Mitochondria Can Produce
Density describes the quantity of mitochondria available. Mitochondrial efficiency describes how productively they operate. Mitochondrial efficiency refers to how much ATP is produced per unit of oxygen consumed — a ratio that is not fixed and changes with training status.
At submaximal aerobic intensities, well-trained mitochondria preferentially oxidize fat as a primary substrate. Fat oxidation yields more total ATP per molecule than carbohydrate oxidation, but it requires a higher rate of oxygen delivery and is only feasible when mitochondrial enzyme density is sufficient to process it at the necessary rate. This is the cellular explanation for why trained athletes burn a higher proportion of fat at any given absolute workload: their fat oxidation pathway more completely supported by mitochondrial enzyme capacity (Romijn et al. 1993). Athletes with lower mitochondrial density reach their carbohydrate-dependent threshold earlier, at lower absolute intensities.
Within mitochondria, the oxidative enzymes that drive aerobic metabolism, specifically citrate synthase, succinate dehydrogenase, and the beta-oxidation enzymes, increase in concentration with aerobic training (Holloszy 1967). Higher enzyme density means the mitochondrion can process substrate more completely and at higher rates. The practical consequence is that two athletes at the same heart rate and the same external power output can be working at very different percentages of their mitochondrial capacity, with significantly different amounts of headroom remaining before fatigue accumulates.
KEY AEROBIC ENZYMES
Citrate Synthase
Entry-point enzyme of the citric acid cycle. Its concentration in muscle biopsies is a standard proxy for mitochondrial density.
Succinate Dehydrogenase
Links the citric acid cycle to the electron transport chain. Upregulated with aerobic training.
Beta-Oxidation Enzymes
Drive fat breakdown inside the mitochondrion. Increased activity enables higher fat oxidation rates at aerobic intensities.
What Is Mitochondrial Biogenesis: How the Body Builds More Mitochondria
Mitochondrial density is not a fixed characteristic. The biological process by which cells increase their mitochondrial mass is called mitochondrial biogenesis, and it involves the synthesis of new mitochondrial proteins and the replication of mitochondrial DNA within existing cells (Hood 2001).
The primary molecular regulator of this process is a protein called PGC-1alpha. When activated, PGC-1alpha triggers a downstream gene expression cascade that increases the synthesis of mitochondrial proteins and upregulates oxidative capacity in the trained muscle (Puigserver and Spiegelman 2003). Exercise is the most reliably documented activator of PGC-1alpha in skeletal muscle, with upregulation detectable after a single bout of endurance work (Baar et al. 2002).
Two distinct cellular signals activate PGC-1alpha through different upstream pathways. The first is AMPK, an energy-sensing enzyme that becomes activated when cellular ATP levels fall relative to AMP, which occurs during sustained low-intensity aerobic work. The second is calcium signaling, which is triggered by high-frequency muscle contractions characteristic of higher-intensity effort. These two pathways operate independently and are additive: activating one does not substitute for the other, which is why different types of training produce different contributions to biogenesis (Coffey and Hawley 2007).
This is why mitochondrial density functions as a performance variable independent of cardiovascular fitness: it sets the oxidative ceiling within the muscle itself.
Biogenesis is a slow process. Initial adaptations, primarily enzyme upregulation within existing mitochondria, can appear within two to four weeks of consistent appropriate training. Meaningful increases in actual mitochondrial mass take four to eight weeks or longer, and full structural adaptation develops over months of sustained stimulus (Hood 2001). One mechanistic aside relevant to supplementation: cordycepin, a nucleoside derived from cordyceps mushrooms, has been shown in cell and animal models to modulate AMPK signaling. The evidence in exercising humans is preliminary and is examined separately in the companion article on cordyceps and mitochondrial function.
These two pathways operate independently and are additive: activating one does not substitute for the other, which is why different types of training produce different contributions to biogenesis.
What Mitochondrial Density Means for Performance Markers You Actually Track
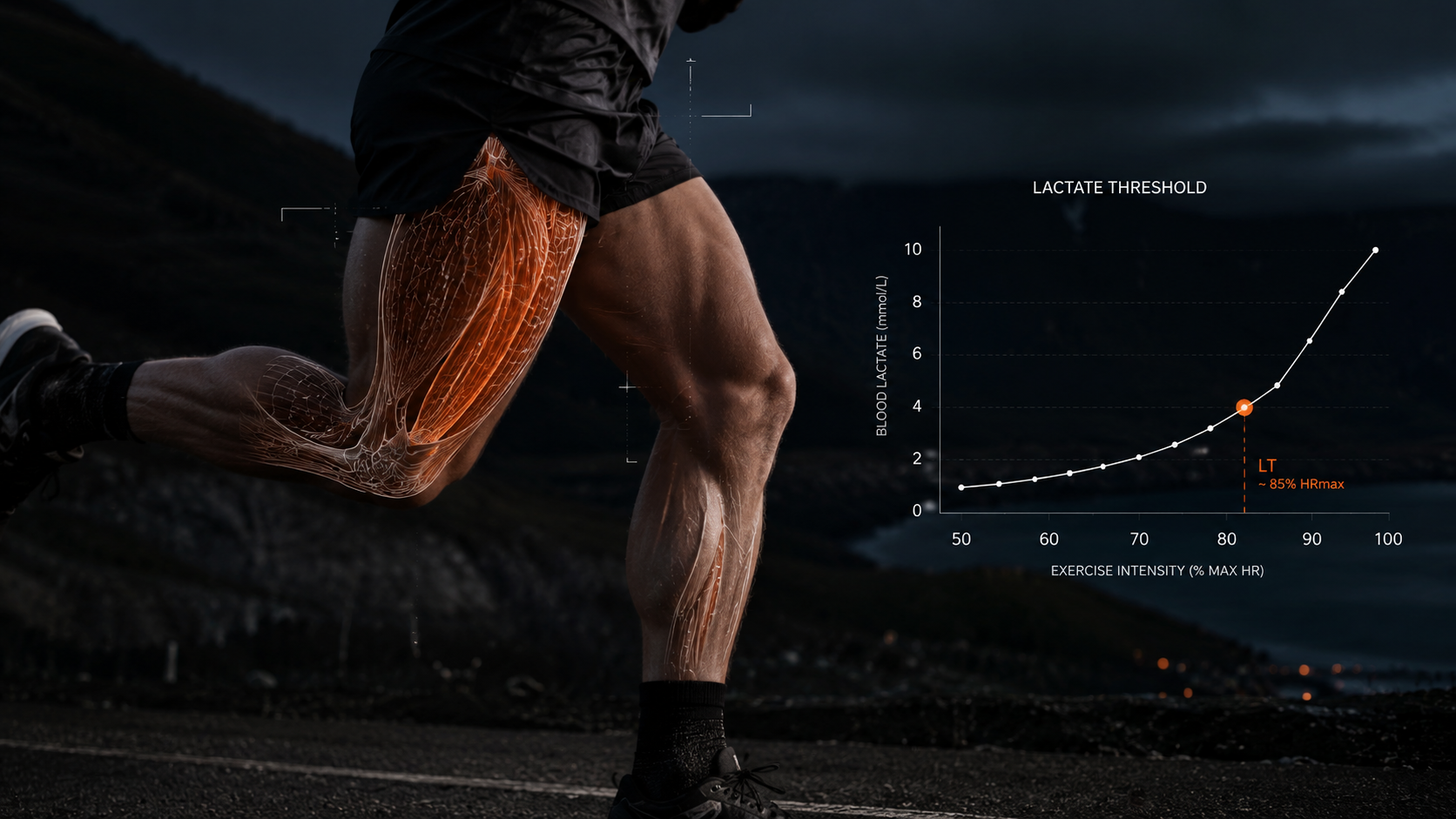
The relationship between mitochondrial density and performance is not abstract. It expresses itself directly in the metrics athletes measure and train around. Lactate threshold is the exercise intensity at which lactate accumulates in the blood faster than it can be cleared. The cellular mechanism behind this threshold is largely mitochondrial. Glycolysis produces pyruvate as its end product. When mitochondria have sufficient capacity, pyruvate enters the mitochondria and is oxidized aerobically. When the work rate exceeds mitochondrial processing capacity, pyruvate accumulates faster than it can be absorbed and is converted to lactate instead. Higher mitochondrial density raises the absolute work rate at which this shift begins (Holloszy and Coyle 1984). Training that improves lactate threshold does so primarily by increasing mitochondrial capacity. The performance marker and the underlying mechanism are causally connected, not merely correlated.
LOW MITOCHONDRIAL DENSITY
Lactate Threshold Reached Earlier
Pyruvate exceeds mitochondrial processing capacity at lower intensities. Glycogen depletes faster. Sustainable power ceiling is lower.
HIGH MITOCHONDRIAL DENSITY
Extended Aerobic Range
More pyruvate processed aerobically before lactate accumulates. Higher fat oxidation rate. Sustainable power ceiling is meaningfully higher.
Fat oxidation rate at a given absolute workload is also a direct expression of mitochondrial capacity. An athlete who can sustain a higher proportion of energy demand from fat at aerobic intensities preserves glycogen stores, delaying the power loss associated with carbohydrate depletion. This reflects the density and enzyme activity of mitochondria in the working muscle, and it can be measured through fat max testing, the intensity at which fat oxidation peaks (Romijn et al. 1993).
The capacity to hold a given power output without progressive decline is also shaped by how far below mitochondrial ceiling the athlete is working. An athlete with low mitochondrial density reaches that ceiling at a low absolute workload. An athlete with high mitochondrial density has more margin before glycolytic reliance increases, lactate accumulates, and contractile capacity begins to drop. Lactate threshold testing and respiratory exchange ratio assessments at submaximal intensities both provide indirect reads on mitochondrial capacity that are more mechanistically informative than VO2max alone.
Lactate threshold testing and respiratory exchange ratio assessments at submaximal intensities both provide indirect reads on mitochondrial capacity that are more mechanistically informative than VO2max alone.
Why Training Harder Doesn’t Always Build More Mitochondria

The most common training error in this context follows directly from conflating VO2max with mitochondrial density.
High-intensity training reliably improves VO2max, primarily through cardiac adaptations: increased stroke volume, higher cardiac output, and greater peak oxygen delivery. These are real and important adaptations. But cardiac remodeling can raise VO2max substantially without producing proportional increases in mitochondrial density in skeletal muscle (Coffey and Hawley 2007). An athlete who raises their VO2max through predominantly high-intensity work may have improved the delivery side of the equation while leaving the utilization side largely unchanged. The cardiovascular ceiling rises; the mitochondrial ceiling does not follow automatically.
The second error is accumulating most training volume in moderate-intensity zones. This range feels productive: heart rate is elevated, sessions generate fatigue, and effort seems substantial. The problem is that moderate-intensity work does not strongly activate either biogenesis pathway. It is not sustained and low enough to generate maximal AMPK activation, and it is not intense enough to produce the calcium signaling associated with high-frequency contractions. Research on training intensity distribution in elite endurance athletes consistently shows that superior aerobic adaptations follow a polarized distribution: approximately 75 to 80 percent of training at low intensity and a smaller amount at true high intensity, with minimal time in the moderate zone (Seiler 2010). Athletes who train predominantly in the middle accumulate fatigue without maximizing either adaptation pathway.
4.1
Training Only for VO2max
Cardiac adaptations improve oxygen delivery but don't proportionally increase mitochondrial density. The utilization ceiling stays where it was.
4.2
Training Primarily in the Moderate Zone
Neither the AMPK pathway nor the calcium pathway is strongly activated. Fatigue accumulates; biogenesis signal remains weak.
4.3
Abandoning Volume Before Adaptation
Mitochondrial biogenesis takes 4 to 8 weeks or longer of consistent stimulus. Switching to harder training before structural changes occur resets the adaptation clock.
When athletes shift to higher aerobic volume and do not observe rapid performance changes, they often return to harder, more immediately fatiguing training. Mitochondrial biogenesis requires weeks of consistent stimulus before structural changes become measurable. Discontinuing the protocol during the pre-adaptation window is a mechanistic error: the biogenesis signal is present, but the structural result never materializes.
Discontinuing the protocol during the pre-adaptation window is a mechanistic error: the biogenesis signal is present, but the structural result never materializes.
Building Mitochondrial Density: What the Mechanism Prescribes

Why Zone 2 Is the Most Efficient Mitochondrial Stimulus
Zone 2 describes training at an intensity where fat oxidation is near its maximum and sustained conversation is possible but effortful. At this intensity, sustained AMPK activation occurs without the glycolytic demand that accompanies higher intensities. Because AMPK activation is the upstream trigger for PGC-1alpha and biogenesis, this intensity range generates a potent mitochondrial adaptation signal while imposing an accumulated metabolic cost low enough to allow high weekly training volume. The aerobic enzymes that enable fat oxidation are themselves upregulated through this same pathway, which is why fat oxidation capacity and zone 2 adaptation develop together over time.
Volume drives cumulative adaptation. A single session creates the biogenesis signal, but structural mitochondrial changes require repeated exposures over weeks. The value of zone 2 training is not any individual session; it is the capacity to accumulate high volumes of stimulus at a training cost that makes that accumulation sustainable. Athletes who want the full protocol rationale will find it covered in the companion article on zone 2 and mitochondrial adaptation.
How to Increase Mitochondria: The Structural Logic of Aerobic Training
Because mitochondrial biogenesis is driven by two distinct signaling pathways, a well-constructed aerobic training program includes both types of stimulus. Low-intensity volume addresses the AMPK pathway. High-intensity intervals address the calcium signaling pathway. Both contribute, neither fully substitutes for the other, and optimal programs include both in appropriate proportion (Coffey and Hawley 2007; Gibala et al. 2006).
AMPK PATHWAY
Activated by Sustained Low-Intensity Work
Energy depletion triggers AMPK, which activates PGC-1alpha and mitochondrial protein synthesis. The primary driver of density gains from aerobic volume.
CALCIUM PATHWAY
Activated by High-Frequency Contractions
High-intensity effort triggers calcium release, activating a parallel biogenesis signal. Additive with AMPK, not interchangeable with it.
For athletes who are currently undertrained in aerobic volume, adding low-intensity work is the higher-return intervention. The limiting factor is not intensity; it is aerobic base volume, which is the stimulus that most directly drives mitochondrial biogenesis through the AMPK pathway. The core structural principle is this: adaptation occurs when training consistently demands more oxidative output than the current mitochondrial supply can comfortably meet. Recovery is load-bearing in this system. Biogenesis happens during rest, not during the training session.
Programs that generate high systemic fatigue without sufficient rest between sessions create the biogenesis signal without allowing protein synthesis to complete.
Which Exercises Build Mitochondria Most Effectively
Any sustained aerobic exercise that engages large muscle groups at appropriate intensity and duration generates a mitochondrial biogenesis signal. Running, cycling, rowing, and swimming all produce comparable molecular adaptations when intensity and duration are matched. The exercise modality matters less than the stimulus parameters.
5.1
Large-Muscle Aerobic Work
Running, cycling, rowing, and swimming all generate equivalent biogenesis signals when intensity and duration are matched. Mode matters less than the stimulus.
5.2
Sport-Specific Conditioning
Mitochondrial adaptation is local. Training the exact muscles used in your sport at low intensity produces the most transferable gains.
5.3
Resistance Training
Produces modest mitochondrial adaptation in Type II fibers at higher volumes. A useful complement to aerobic training, not a replacement for it.
What does matter substantially is specificity. Mitochondrial adaptations are largely local to the muscles being trained. Building mitochondrial density in the quadriceps through cycling does not transfer meaningfully to the posterior chain and hip musculature recruited in rowing or in grappling. For athletes whose primary sport is grappling, boxing, or Muay Thai, sport-specific conditioning performed at low intensity produces the most directly transferable mitochondrial adaptation. General aerobic work complements this but does not replicate it.
Resistance training does produce mitochondrial adaptation in Type II fibers, but the biogenesis stimulus is weaker than continuous aerobic work at matched duration. It is a useful supplement to aerobic training, not a functional equivalent.
How Long Does It Take to Increase Mitochondria?
Mitochondrial adaptation unfolds in two overlapping stages. In the first two to four weeks of consistent appropriate training, enzyme concentrations within existing mitochondria increase, raising oxidative capacity before new mitochondria are synthesized (Hood 2001). Some performance benefits appear during this stage because oxidative capacity improves even before structural changes in mitochondrial mass are measurable.
Mitochondrial adaptations are largely local to the muscles being trained.
Actual increases in mitochondrial density take four to eight weeks or longer of consistent training to become detectable, and full adaptation across a training block develops over months. Attempting to accelerate this timeline by increasing training intensity typically counterproduces: higher intensity shortens the available rest window, and biogenesis requires that window to complete. The timeline is not a motivational observation. It is a constraint that follows directly from the biology of protein synthesis and organelle replication.
The timeline is not a motivational observation. It is a constraint that follows directly from the biology of protein synthesis and organelle replication.
Frequently Asked Questions
How long does it take to increase mitochondrial density?
Initial enzyme adaptations within existing mitochondria appear within two to four weeks of consistent aerobic training. Actual increases in mitochondrial mass take four to eight weeks or longer. Full structural adaptation develops over months, which is why aerobic volume programs require sustained commitment before performance changes become measurable.
Does Zone 2 training improve mitochondria?
Zone 2 training produces sustained AMPK activation in skeletal muscle, which is the primary upstream signal for PGC-1alpha and mitochondrial biogenesis. It also promotes fat oxidation enzyme upregulation. Because Zone 2 imposes low accumulated metabolic fatigue, it allows high weekly training volume, which drives cumulative mitochondrial adaptation over time.
What is the difference between VO2max and mitochondrial capacity?
VO2max measures how much oxygen the cardiovascular system can deliver to working muscle. Mitochondrial capacity measures how effectively working muscle uses that delivered oxygen. Both limit performance, but through different mechanisms, and it is possible to improve VO2max through cardiac adaptations without proportionally improving mitochondrial density.
Why do muscle cells need mitochondria?
Muscle cells require mitochondria because sustained mechanical work depends on aerobic ATP production. The anaerobic pathways that produce ATP without oxygen are fast but capacity-limited. For any effort lasting more than a few seconds, oxidative phosphorylation inside mitochondria is the primary energy source. More mitochondria per unit of muscle means more aerobic ATP production capacity at a given moment.
Can resistance training increase mitochondria?
Resistance training does produce mitochondrial adaptation in Type II muscle fibers, particularly at higher training volumes. The biogenesis stimulus is weaker than that from continuous aerobic exercise at matched duration. Resistance training is a useful complement to aerobic training, but it does not replace aerobic work as the primary driver of mitochondrial density.
Performance Capacity Is a Mitochondrial Problem

Aerobic fitness contains two distinct variables that are commonly measured as one. The first is the cardiovascular system’s capacity to deliver oxygen to working muscle. The second is working muscle’s capacity to use that oxygen to produce ATP. Mitochondrial density in skeletal muscle is the primary structural determinant of the second variable, and it responds to different training stimuli and adapts on a different timeline than the first.
Athletes who train primarily to improve cardiovascular output are addressing one half of the system. The other half requires sustained aerobic volume, appropriate intensity distribution, and the rest time for biogenesis to proceed. These two halves of aerobic fitness can diverge substantially in athletes whose training history has emphasized intensity over volume.
The practical reframe is this: when an athlete performs aerobic work at low intensity, the primary adaptation occurring at the cellular level is mitochondrial biogenesis. The cardiovascular benefits co-occur and are real, but the mechanism operating most directly inside the muscle cell is the synthesis of new mitochondrial proteins in response to an energy demand signal. Understanding this changes how a person weighs slow sessions, aerobic base building, and the structure of a training year, because it replaces a vague notion of building fitness with an accurate model of what the training stimulus is actually telling the cell to do.
When an athlete performs aerobic work at low intensity, the primary adaptation occurring at the cellular level is mitochondrial biogenesis.
Your aerobic ceiling is not set by your heart. It is set by how many mitochondria are inside your muscle cells. Consistent aerobic volume, distributed correctly and followed by enough recovery, is what raises it.
Mitochondrial biogenesis takes weeks of repeated stimulus to produce structural change. The slow sessions are not filler; they are the primary signal. The adaptation completes during recovery, not during the session.
Mydos Performance
— What Actually Limits Performance Series —
Mitochondrial Density and Performance Capacity
The cellular machinery behind aerobic capacity, and what actually builds it
By Ricardo Londono, MD/PhD · 2026