
The Problem with How Most People Think About Energy

Most busy successful adults treat energy as something that can be topped up on demand. A cup of coffee in the morning, another at noon, and a pre-workout before training — this is the standard operating model for most of us when managing a full schedule. The problem is that although stimulants produce increased feelings of energy, this is not the same thing as real energy production, and solving a production problem with a perception intervention does not close the gap; it actually defers it.
Instead of trying to take shortcuts to feel more alert, the more important issue is how to improve mitochondrial function to produce more ATP, the molecule that powers every muscular contraction and cognitive process in the body. Caffeine and other stimulants do not increase ATP output (Fredholm et al. 1995). They alter the nervous system's sensitivity to the signals that indicate fatigue has accumulated. The output ceiling remains exactly where it was and has always been.
The consequence of this misalignment is familiar: performance that holds for a while, then drops sharply; reliance on stimulants that increases over time as tolerance builds; a persistent gap between how a person wants to perform and how they actually perform when the caffeine wears off. None of these are caffeine problems. They are capacity problems being bypassed, rather than being addressed. The mechanics of what that bypass actually costs are covered in the article on the caffeine crash.
Perceived Energy
Output Ceiling Unchanged
Stimulants alter the nervous system's sensitivity to fatigue signals. The output ceiling is unchanged.
Actual Energy Production
ATP Synthesis Rate
ATP synthesis rate is determined by mitochondrial capacity. This is the variable that determines repeatable output.
This article is about the system that actually determines how much energy the body can produce: the mitochondria. Specifically, it covers what mitochondria do, what limits their output, how that capacity is built, and what it means to improve mitochondrial function over time rather than managing the perception of it as a shortcut.
Caffeine and other stimulants do not increase ATP output. They alter the nervous system's sensitivity to the signals that indicate fatigue has accumulated.
What Mitochondria Actually Do
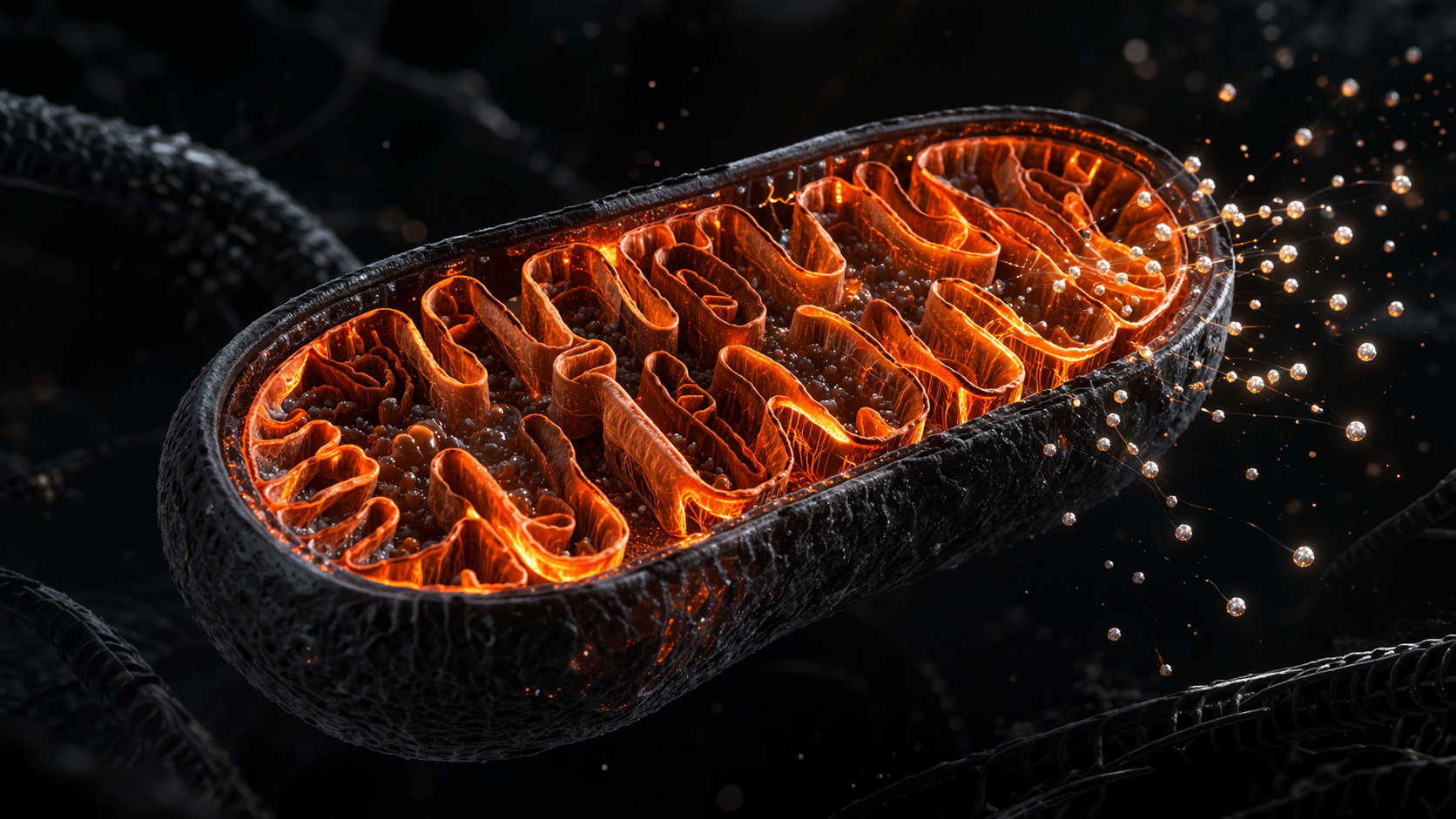
Mitochondria are organelles present in nearly every cell in the human body. Their primary function is converting fuel — carbohydrates, fats, and to a lesser extent proteins — into ATP, using oxygen as the terminal electron acceptor. This process is called oxidative phosphorylation, and it takes place across the inner mitochondrial membrane through a series of protein complexes known collectively as the electron transport chain.
The mechanics are worth understanding at least in outline. As fuel molecules are broken down, electrons are extracted and passed through the transport chain. This movement generates a proton gradient across the inner membrane — a kind of stored electrochemical potential. ATP synthase, an enzyme embedded in that membrane, uses the flow of protons back across it to drive the synthesis of ATP from ADP and inorganic phosphate. The energy that powers a six-minute round of sparring, or a full day of focused work, originates in this membrane-level chemistry.
The efficiency differential between aerobic and anaerobic energy production is substantial. Oxidative phosphorylation yields approximately 30 to 32 ATP molecules per glucose molecule under cellular conditions (Rich et al. 2010). Anaerobic glycolysis — the pathway the body uses when oxygen supply or mitochondrial capacity is insufficient — yields 2 net ATP per glucose. The aerobic pathway is not marginally more efficient. It is an order of magnitude more efficient, which is why aerobic capacity determines how long and how hard a person can sustain output, not whether they can spike it briefly.
30 to 32
ATP per glucose — aerobic pathway
2
Net ATP per glucose — anaerobic glycolysis
15x
Efficiency advantage of aerobic over anaerobic
ATP Yield per Glucose Molecule
31 ATP
Aerobic
(oxidative phosphorylation)
2 ATP
Anaerobic
(glycolysis)
A roughly 15× efficiency gap. Source: Rich et al. 2010 (PMID 21258145).
The rate at which ATP can be produced via oxidative phosphorylation depends on three variables: the number of mitochondria available (density), the efficiency of the electron transport chain within each mitochondrion, and the rate at which oxygen is delivered to the cell. When physical or cognitive demand exceeds what the aerobic system can supply, the body compensates by recruiting anaerobic pathways. Lactate accumulates as a byproduct — not as a toxin, but as a marker that glycolysis is running faster than the mitochondria can process its output (Faude et al. 2009). Performance at that point becomes time-limited rather than sustained.
The aerobic pathway is not marginally more efficient. It is an order of magnitude more efficient, which is why aerobic capacity determines how long and how hard a person can sustain output, not whether they can spike it briefly.
This shift is not a malfunction. It is what happens when demand exceeds aerobic capacity. But it has consequences: lactate accumulation accelerates the subjective fatigue signal, driven by lactate accumulation and afferent nerve signaling from the working muscle, recovery takes longer, and the ability to repeat high-quality efforts diminishes. The ceiling on sustainable work is set by the mitochondria — the level at which a person can perform indefinitely without accumulating unrecoverable fatigue.
The ceiling on sustainable work, the level at which a person can perform indefinitely without accumulating unrecoverable fatigue, is set by the mitochondria.
Why Mitochondrial Density Sets Your Performance Ceiling

Mitochondrial density refers to the number of mitochondria per unit of muscle tissue. Greater density means more parallel sites for ATP synthesis — more throughput from the same fuel and oxygen supply. The analogy that holds is not a faster drip but a wider pipe: the system does not move faster, it moves more.
Two athletes training at identical loads, with identical nutrition and sleep, can have meaningfully different sustained output capacity if their mitochondrial density differs. One person can hold a pace for two hours that exhausts the other in forty minutes. This is not a function of willpower or pain tolerance. It is a function of how many ATP synthesis sites are available in the working muscle, and that variable is measurable (Holloszy et al. 1967).
Sustained Output at Equal Training Load
High mitochondrial density
~2 hrs
Low mitochondrial density
~40 min
Illustrative comparison of the example in the text — same load, nutrition, and sleep.
Density is not fixed. It responds to training, declines with inactivity, and degrades gradually with age in the absence of a sustained aerobic stimulus. An athlete who trains consistently for years accumulates mitochondrial density. An athlete who takes three months off loses a portion of it. This is not a metaphor for fitness. It is a structural change in the muscle tissue, measurable through biopsy and reflected in performance.
What Changes Mitochondrial Density
Training
Sustained aerobic stimulus activates the biogenesis pathway, producing structural increases in mitochondrial density over weeks to months.
Inactivity
Sedentary periods reduce mitochondrial density as the adaptive stimulus is removed and existing mitochondria degrade.
Age
Mitochondrial density declines gradually with age in the absence of a consistent aerobic stimulus.
Training that prevents adequate recovery interrupts the biogenesis cycle without producing the density increase.
Understanding density reframes what it means to improve mitochondrial function. The goal is not to make existing mitochondria operate faster in the short term. Mitochondria are already operating near their enzymatic limits during high-intensity work. The goal is to build more of them, maintain their efficiency, and protect the conditions that allow the adaptation to compound over time. Anything that interferes with that process — chronic overtraining, inadequate recovery, or dependency on stimulants that mask fatigue signals and lead to underrecovery — reduces the ceiling rather than raising it.
Two athletes training at identical loads, with identical nutrition and sleep, can have meaningfully different sustained output capacity if their mitochondrial density differs.
How the Body Builds More Mitochondria

Mitochondrial biogenesis is the process by which cells synthesize new mitochondria. It is not a continuous background process. It is triggered by a specific signal: the detection of energy demand that exceeds what the current mitochondrial population can efficiently meet. Sustained aerobic exercise creates exactly this signal, and the cellular machinery that responds to it is now well characterized.
The primary driver is a protein called PGC-1α — peroxisome proliferator-activated receptor gamma coactivator 1-alpha. PGC-1α functions as a transcriptional coactivator: when activated, it binds to and enhances the activity of transcription factors that control the expression of genes encoding mitochondrial proteins (Puigserver et al. 2003). In practical terms, PGC-1α activation is the upstream signal that initiates the construction of new mitochondria. It is not the mitochondria themselves. It is the instruction to build them.
PGC-1α is activated by sustained aerobic work, particularly through two upstream signals: AMPK (AMP-activated protein kinase), which detects low cellular energy charge, and SIRT1, a deacetylase that responds to NAD+ availability [CITE: AMPK PGC-1alpha exercise intensity mitochondrial biogenesis — PMID 23287861 requires manual PubMed verification before inserting]. Both of these signals increase during sustained moderate-intensity exercise and decline rapidly at rest. The implication is that the biogenesis signal is dose-dependent and accumulates with training duration, not just intensity.
4.1
The Signal
Sustained aerobic work activates AMPK and SIRT1, which upregulate PGC-1α. This is the upstream instruction to build new mitochondria.
4.2
The Timeline
A single session activates the pathway but produces no structural change. Density increases require weeks to months of repeated activation.
4.3
The Recovery Window
Protein synthesis that completes biogenesis happens during rest. Training that prevents recovery interrupts the cycle before the density gain is captured.
The Biogenesis Signal Cascade
Stimulants do not enter this cascade — they activate none of these signals.
The timeline for biogenesis is not acute. Measurable increases in mitochondrial density require weeks to months of consistent training stimulus. A single session activates PGC-1α but does not produce structural change. The structural change accumulates through repeated activation across many sessions, provided that recovery between sessions is sufficient to allow protein synthesis to complete. Training that prevents adequate recovery interrupts this cycle without producing the density increase that justified the stress in the first place.
PGC-1α activation is the upstream signal that initiates the construction of new mitochondria. It is not the mitochondria themselves. It is the instruction to build them.
The stimulus must also be appropriately calibrated. Work that is too easy does not generate sufficient AMPK activation or energy stress to robustly trigger PGC-1α. Work that is too hard produces inflammatory and hormonal signals that compete with or suppress the biogenesis pathway. The intensity range that reliably drives mitochondrial adaptation without compromising recovery is not arbitrary. It corresponds to what sports science classifies as moderate aerobic intensity, often called Zone 2.
This is the physiological basis for a point that is easy to state but frequently ignored: caffeine and other stimulants do not touch the biogenesis pathway. They do not activate AMPK. They do not upregulate PGC-1α. They create no signal that the cell interprets as a reason to build more mitochondria. Using stimulants as an energy management strategy is orthogonal to the process that actually improves energy production capacity.
Caffeine and other stimulants do not touch the biogenesis pathway. They do not activate AMPK. They do not upregulate PGC-1α.
Why Stimulants Borrow Energy Instead of Building It
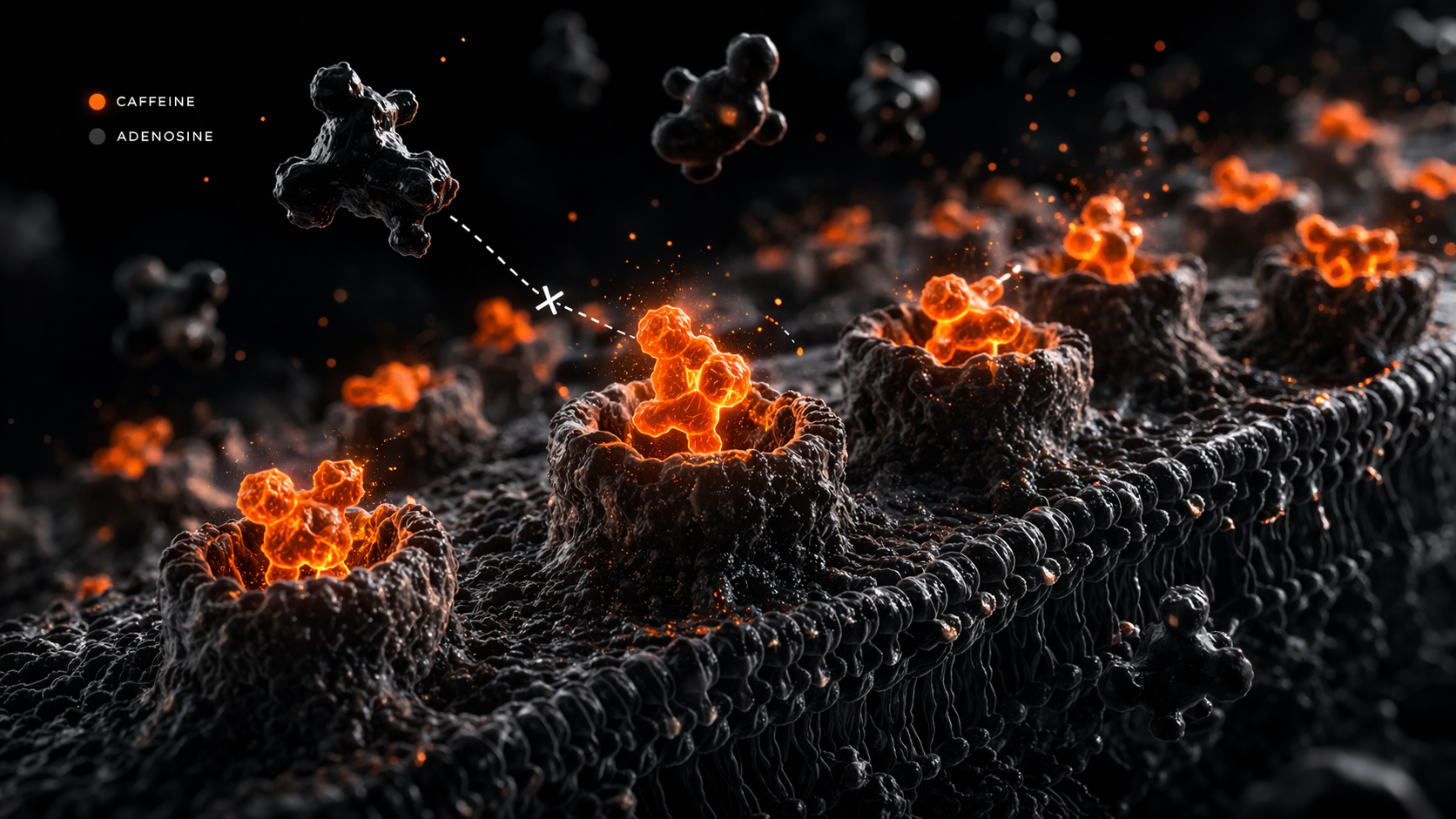
A caffeine crash is not a side effect. It is the mechanism working exactly as designed, arriving at the predictable end of its cycle. Understanding what caffeine actually does at the receptor level makes this clear and makes the crash legible rather than mysterious.
Adenosine is a byproduct of cellular activity that accumulates in the brain and body during wakefulness. As it accumulates, it binds to adenosine receptors — particularly A1 and A2A receptors — and produces the subjective experience of fatigue and the drive to sleep. This is a regulatory system, not a malfunction: adenosine signals that metabolic work has been done and that rest is warranted — the fatigue signal implicated in both central and peripheral fatigue pathways. Caffeine is a adenosine receptor antagonism — a competitive antagonist of these receptors (Fredholm et al. 1995). It occupies the receptor sites without activating them, blocking adenosine from binding, and alertness is maintained.
Three things do not change during caffeine's action: ATP production rate, substrate availability, and mitochondrial function. The output ceiling is identical before and after caffeine. What shifts is the perception of proximity to that ceiling — the point at which fatigue registers consciously. A person under caffeine's effect can continue working at a level that would otherwise feel unsustainable, but the metabolic cost of that work is unchanged. The fatigue is not removed; it is deferred.
What Caffeine Changes
Fatigue Perception
Blocks adenosine receptors. Prevents the fatigue signal from completing its pathway. Perception of proximity to the output ceiling shifts upward.
What Caffeine Does Not Change
The Output Ceiling Itself
ATP production rate. Substrate availability. Mitochondrial function. The output ceiling itself remains exactly where it was.
The caffeine crash is what happens when caffeine is metabolized and adenosine receptor blockade ends (Nehlig et al. 1992). During the period of blockade, adenosine has continued to accumulate. When the receptors become available again, that accumulated adenosine binds rapidly and the deferred adenosine load presents itself in force. This is not a blood glucose crash, and it is not caused by caffeine itself wearing off in some general sense. The severity of the crash correlates with how much adenosine accumulated during the suppression period, which is a function of how much metabolic work was done and how long the caffeine was active.
The fatigue is not removed; it is deferred.
Habitual caffeine use does not improve the underlying system. Adenosine receptor density upregulates in response to chronic blockade, meaning the body compensates by producing more receptors, requiring more caffeine to achieve the same suppressive effect. Some evidence suggests that chronic high-dose caffeine intake may affect mitochondrial efficiency, though this evidence is not conclusive and should not be overstated. What is clear is that the stimulant cycle — caffeine to perform, crash to recover, caffeine again — accumulates fatigue across sessions without generating the aerobic adaptation that would reduce the gap being managed.
The stimulant cycle — caffeine to perform, crash to recover, caffeine again — accumulates fatigue across sessions without generating the aerobic adaptation that would reduce the gap being managed.
What a Caffeine Crash Reveals About Your Energy System

Caffeine crash symptoms — fatigue, reduced concentration, irritability, and low motivation — are not random or idiosyncratic. They are the predictable result of adenosine receptor blockade ending and deferred fatigue signal arriving. But beyond explaining the mechanism, the severity and frequency of these symptoms carry diagnostic information about the state of the underlying energy system.
A person with high mitochondrial capacity has a higher baseline ATP production rate. Their aerobic system covers a larger portion of their daily energy demand without requiring anaerobic supplementation or stimulant masking. When caffeine wears off, the gap between stimulated state and baseline state is smaller, because the baseline is higher. The crash is less severe — not because of anything caffeine-specific, but because there is less distance to fall.
Frequent or severe crashes point to a baseline energy system operating below demand. The caffeine was not enhancing performance so much as bridging a gap that existed independent of it. That gap — the difference between what the system can produce and what daily functioning requires — is primarily an aerobic capacity gap. Addressing it with better caffeine timing or switching to a different stimulant does not close the gap. It manages the symptoms of a gap that remains open.
High Mitochondrial Capacity
Smaller Gap, Milder Crash
Aerobic system covers a larger portion of daily energy demand. When caffeine wears off, the gap between stimulated and baseline state is smaller. The crash is less severe.
Low Mitochondrial Capacity
Larger Gap, Harder Crash
Aerobic system operates below daily demand. Caffeine bridges the gap rather than enhancing performance. When it wears off, the gap is larger and the crash is more severe.
The question to ask after a bad crash is not what to take next time. The question is what the crash indicates about the current state of aerobic capacity and whether the training and recovery inputs being applied are sufficient to move that capacity in the right direction.
Frequent or severe crashes point to a baseline energy system operating below demand.
Training and Recovery as Capacity Builders

Mitochondrial biogenesis is primarily driven by sustained aerobic work at moderate intensity. In the framework used by exercise physiologists, this corresponds to Zone 2: output below the lactate threshold, where the aerobic system is stressed consistently without being overwhelmed. At this intensity, the metabolic signals that activate PGC-1α — particularly AMPK activation from low cellular energy charge — are generated and sustained for the duration of the session without producing the inflammatory and catabolic signals associated with high-intensity work (Iaia et al. 2008).
The training dose that produces mitochondrial adaptation is not trivial. Research on endurance training and aerobic adaptation points to sustained sessions of 45 to 90 minutes at Zone 2 intensity, repeated three to four times per week, maintained across weeks and months (Stoggl et al. 2014). This duration reflects the time required to generate sufficient PGC-1α activation and mitochondrial protein synthesis signals. Shorter sessions at this intensity may produce some signal, but the evidence for meaningful structural adaptation comes from sustained aerobic volume. The specific numbers carry uncertainty — the threshold is not a fixed line — but the general principle is well-supported: mitochondrial adaptation is a volume-dependent process, not an intensity-dependent one.
Zone 2 Training — The Biogenesis Stimulus
Intensity
Output below the lactate threshold. Aerobic system stressed consistently without being overwhelmed. AMPK activated without generating excessive inflammatory signals.
Duration
Sessions of 45 to 90 minutes. Sufficient time for PGC-1α activation and mitochondrial protein synthesis signals to accumulate.
Frequency
Three to four sessions per week. Repeated stimulus required for structural density change — not a single-session effect.
Recovery
Sleep is the window for protein synthesis. Inadequate recovery interrupts the biogenesis cycle before the adaptation is captured.
Mitochondrial Density Builds Over Weeks, Not Sessions
Illustrative trajectory. Adaptation is volume-dependent and accumulates across many sessions with adequate recovery.
The implication for someone who trains combat sports or strength work alongside aerobic conditioning is worth stating plainly. High-intensity sessions — sparring rounds, heavy lifting, interval work — produce different and in some ways competing adaptations. They are not substitutes for Zone 2 in terms of mitochondrial biogenesis. A training week that consists entirely of high-intensity work without sustained aerobic sessions generates limited PGC-1α signaling, regardless of how hard the sessions feel, because the aerobic system requires aerobic stimulus to adapt. The interaction between high-intensity and aerobic training across a week is the subject of the fitness-fatigue model.
Mitochondrial adaptation is a volume-dependent process, not an intensity-dependent one.
Recovery is the other half of the equation, and it is not passive. The synthesis of mitochondrial proteins following exercise occurs primarily during sleep, when growth hormone secretion peaks and catabolic signaling from cortisol is suppressed [CITE: sleep mitochondrial protein synthesis growth hormone — PMID 21550729 is Medical Hypotheses tier; verify or soften GH claim if stronger citation unavailable]. Inadequate sleep does not merely blunt the feeling of recovery. It limits the biological process through which training translates into structural adaptation. The mitochondrial density gains that were triggered by the training session require the recovery environment to be completed.
The practical framing for how to improve mitochondrial function naturally is therefore not a list of interventions. It is a coherent system: consistent aerobic stimulus at the right intensity, sufficient volume over time, and recovery conditions that allow the adaptation to complete. These variables are not independent — chronic sleep deprivation impairs training quality, and inadequate aerobic volume limits how much high-intensity work can be supported — but each is necessary. Stimulants do not substitute for any of them.
The practical framing for how to improve mitochondrial function naturally is therefore not a list of interventions. It is a coherent system: consistent aerobic stimulus at the right intensity, sufficient volume over time, and recovery conditions that allow the adaptation to complete.
Where Cordyceps Fits in the Capacity Framework

Cordyceps — a fungus with a long history of use in traditional Chinese medicine — has attracted research interest specifically because its proposed mechanism of action is distinct from stimulant-based energy support. The active compounds, including cordycepin (a structural analog of adenosine) and related adenosine derivatives, are thought to interact with adenosine receptors and cellular energy metabolism in ways that may support oxygen utilization rather than mask fatigue signals.
A double-blind, placebo-controlled trial in older adults found that supplementation with a standardized Cordyceps extract improved exercise performance and VO2 max relative to placebo (Chen et al. 2010). The authors proposed enhanced oxygen utilization as the underlying mechanism, though the cellular pathway was not directly measured in that study. The evidence base for cordyceps in healthy, trained adults is limited — most human trials are small, and the mechanistic claims remain proposed rather than confirmed.
The distinction from stimulants is mechanistic rather than incidental. Caffeine works by blocking adenosine receptors and deferring fatigue signals. Cordyceps appears to operate differently — potentially by supporting the efficiency of the aerobic pathway itself rather than altering how the nervous system responds to its output. This is the difference between borrowing from a system and supporting it, though this framing requires the caveat that the supporting evidence is still preliminary.
Stimulants (Caffeine)
Borrows from the System
Competitive antagonist of adenosine receptors. Blocks the fatigue signal. Does not change ATP production rate, substrate availability, or mitochondrial function.
Cordyceps (Proposed)
Supports the System
Proposed to support efficiency of the aerobic pathway itself via cordycepin and adenosine analogs. If the mechanism is real, it acts on the production side. Evidence is preliminary.
Within the framework this article has outlined, cordyceps belongs to a different category than stimulants: it is a support variable that may modulate aerobic function, not one that substitutes for the training and recovery inputs that build aerobic capacity. Whether it produces a measurable effect for a given individual depends on factors the current evidence cannot fully specify. What the evidence does not support is the framing that it provides energy in any way analogous to caffeine. The mechanism, if real, points in the opposite direction.
Cordyceps belongs to a different category than stimulants: it is a support variable that may modulate aerobic function, not one that substitutes for the training and recovery inputs that build aerobic capacity.
Frequently Asked Questions
What does it mean to improve mitochondrial function?
Improving mitochondrial function means increasing the number and efficiency of mitochondria in muscle cells, which raises the ceiling on how much ATP the body can produce aerobically. This happens through sustained aerobic training, primarily Zone 2 work, maintained over weeks and months. No supplement or stimulant substitutes for this process.
Why does caffeine cause a crash?
Caffeine blocks adenosine receptors, preventing the fatigue signal from completing its pathway. Adenosine continues to accumulate while the block is active. When caffeine is metabolized, receptors become available and the accumulated adenosine binds simultaneously, producing the fatigue rebound called the crash. The crash is deferred fatigue arriving, not a separate effect of caffeine itself.
Does caffeine improve mitochondrial function?
No. Caffeine does not activate the signaling pathways that drive mitochondrial biogenesis. It does not increase AMPK activity, upregulate PGC-1α, or produce any signal the cell interprets as a reason to build more mitochondria. Caffeine alters fatigue perception; it does not change the capacity of the system producing energy.
What is mitochondrial density and why does it matter?
Mitochondrial density refers to the number of mitochondria per unit of muscle tissue. Higher density means more sites for ATP synthesis operating in parallel, which increases the amount of aerobic work that can be sustained before the body shifts to less efficient anaerobic pathways. It is the primary determinant of repeatable output capacity.
How long does it take to improve mitochondrial function through training?
Measurable increases in mitochondrial density require weeks to months of consistent aerobic stimulus. A single training session activates the biogenesis pathway but does not produce structural change. The adaptation accumulates through repeated activation across many sessions, provided recovery between sessions is adequate for mitochondrial protein synthesis to complete.
The Bottom Line
Mitochondria set the ceiling on sustainable energy output. Every watt of sustained power, every hour of focused cognitive work, every training session that does not require days to recover from — these are expressions of mitochondrial capacity. Everything above that ceiling is borrowed: from stimulants that defer fatigue signals, from anaerobic reserves that take longer to replenish, from recovery that gets compressed because performance demands did not allow for it.
The ceiling is not fixed. Consistent aerobic training at the right intensity activates the biogenesis pathway — specifically PGC-1α — and over weeks and months, mitochondrial density increases. The ceiling rises. Recovery, particularly sleep, is the window in which the structural changes triggered by training are completed. Without it, the stimulus accumulates but the adaptation does not.
Most energy management strategies — caffeine, pre-workout compounds, energy drinks — operate beneath the ceiling without changing it. They alter the perception of proximity to a limit that remains where it was. Some, through the dependency and underrecovery cycles they enable, may lower the ceiling over time rather than sustaining it. This is not an argument against their use in specific, appropriate contexts. It is an argument for understanding what they actually do, so that a person managing a serious training load alongside real-life demands is solving the right problem.
The people who perform well at 45 and 50 under real load, with full careers and families and serious training, are, in most cases, the people who have been building the system rather than borrowing from it.
Understanding how to improve mitochondrial function is not a route to a small and temporary increment of performance. It is a route to performing at a level that compounds over years rather than declining as the stimulant strategies that were holding things together stop working. The people who perform well at 45 and 50 under real load — with full careers and families and serious training — are, in most cases, the people who have been building the system rather than borrowing from it.
Your mitochondria set the ceiling. Everything else — caffeine, stimulants, pre-workout — masks the feeling.
The ceiling moves, but only through one mechanism: building more mitochondria over time through consistent aerobic training and adequate recovery. There is no acute intervention that substitutes for this process.
Mydos Performance
— Sustained vs Borrowed Energy Series —
How Mitochondria Actually Work
The Science Behind Real Energy Output
By Ricardo Londono, MD/PhD