
The Wall You Did Not See Coming

Hitting the wall running feels different from running out of steam. At mile 20, sometimes earlier at mile 18 and sometimes later at mile 22, something changes categorically: pace collapses, legs lose their response, and the effort required to maintain even a fraction of race speed escalates beyond what feels manageable. The deterioration is abrupt enough that experienced runners describe it as a distinct event, a moment when something breaks.
The phenomenon goes by two names in endurance sport. Hitting the wall refers to the sudden performance collapse associated with marathon-distance running. Bonking in running and cycling describes the same cluster of symptoms, with a specific emphasis on hypoglycemia, meaning a significant drop in blood glucose, as the precipitating cause. In practice, the two overlap substantially. The wall involves both muscular glycogen depletion and blood glucose decline, and the symptoms of each compound the other.
The common interpretation is that the wall reflects mental failure or inadequate preparation: the athlete did not train hard enough, went out too fast, or could not hold on psychologically. These factors are real contributors to the timing of the wall, but they describe when it happens, not what causes it. The mechanism of hitting the wall running is cellular, governed by substrate availability, enzyme kinetics, and the rate at which the body can convert fuel to ATP at a given exercise intensity. Understanding this distinction separates athletes who manage the wall from those caught off guard by it, and it changes what meaningful preparation actually looks like.
COMMON EXPLANATION
What People Attribute the Wall To
The athlete trained insufficiently, went out too fast, or could not hold on psychologically. These factors affect when the wall happens.
WHAT CAUSES IT
What Actually Governs the Wall
The wall is governed by substrate availability, enzyme kinetics, and the rate at which the body can convert fuel to ATP at a given exercise intensity.
The mechanism of hitting the wall running is cellular, governed by substrate availability, enzyme kinetics, and the rate at which the body can convert fuel to ATP at a given exercise intensity.
The Finite Fuel Tank: Glycogen, Depletion, and the Cell

The human body runs on two fuel sources during endurance exercise: carbohydrate and fat. Both are oxidized simultaneously, but the ratio shifts depending on exercise intensity. Above roughly 65 to 70 percent of VO2max, the range that encompasses marathon race pace for most trained runners, carbohydrate becomes the dominant fuel contributor, providing approximately 80 to 90 percent of the energy for ATP resynthesis at high intensities (Romijn et al. 1993). Fat continues to be oxidized in parallel, but its contribution shrinks as intensity climbs.
This intensity-dependence is not a preference the body can override with training or willpower. It is a function of how fast different substrates can be metabolized relative to the rate at which ATP is being consumed.
How Much Glycogen Your Muscles Can Store
The glycogen stored in muscle and liver is the finite fuel that high-intensity aerobic work depletes. Muscle glycogen, the primary fuel source for working muscle fibers, totals approximately 300 to 400 grams in most adults, varying with training status and body size. Liver glycogen adds another 80 to 100 grams. Total carbohydrate stores sit around 400 to 500 grams, equivalent to roughly 1,600 to 2,000 kilocalories (Hawley and Leckey 2015). At marathon pace, muscle glycogen is consumed at approximately 1 gram per minute.
~400–500g
TOTAL GLYCOGEN STORED
~1g/min
DEPLETION RATE AT MARATHON PACE
90–120 min
WINDOW BEFORE CRITICAL DEPLETION
Running the math makes the wall predictable: a runner with 400 grams of stored muscle glycogen, running at race pace from the start, depletes that supply in roughly 90 to 120 minutes. For a 3:30 to 4:00 marathon, that places the window of critical depletion squarely at miles 18 to 22, precisely where the wall is classically observed. Trained athletes and athletes who have used carbohydrate loading protocols can expand this ceiling modestly, but they cannot eliminate it. The fuel tank, regardless of preparation, has a hard limit.
What Glycogen Depletion Running Actually Means
Glycogen depletion running is not simply the condition of having low glycogen. The wall does not arrive gradually as stores decline. Performance holds relatively stable through much of the depletion process, then drops sharply once glycogen falls below a critical threshold, the point at which it can no longer supply ATP fast enough to sustain current workload (Coyle et al. 1986). This non-linear relationship between glycogen availability and performance output is why the wall feels sudden even though depletion is progressive.
Some glycogen remains in the tank when the wall strikes. The fuel is not gone; what is gone is the rate at which that fuel can turn over. Above the threshold intensity, the body's glycogen-derived ATP supply falls short of demand before stores are exhausted. The athlete experiences this as an abrupt ceiling on output, not as a gradual fade into fatigue.
From Glycogen to ATP: The Energy Chain
FROM GLYCOGEN TO ATP
Glycogen → Glucose
Stored glycogen is broken down to glucose inside the muscle cell, making it available for the next step in the energy chain.
Glycolysis → Pyruvate
Glucose enters glycolysis, producing pyruvate and a small initial ATP yield. This step does not require oxygen.
Oxidative Phosphorylation → ATP
Pyruvate enters the Krebs cycle; the bulk of ATP is generated via oxidative phosphorylation in the mitochondria — the primary site of aerobic energy production.
The relevance of glycogen to the wall is mediated through ATP, the molecule that directly powers muscle contraction. Glycogen is broken down to glucose, which enters glycolysis and produces pyruvate, which feeds the Krebs cycle and ultimately generates ATP via oxidative phosphorylation in the mitochondria. The rate at which this chain can turn over determines whether fuel supply matches demand. At high exercise intensities, the demand for ATP resynthesis is high, and carbohydrate-derived substrates meet this demand more efficiently than fat-derived substrates. This is the core of why glycogen is not interchangeable with fat at race pace, and why running out of glycogen produces a performance collapse rather than a smooth fuel transition.
This is the core of why glycogen is not interchangeable with fat at race pace, and why running out of glycogen produces a performance collapse rather than a smooth fuel transition.
Why Fat Oxidation Is Too Slow for Race Pace
The argument against fat as a backup fuel during the marathon wall is frequently misframed as "fat stores run out." They do not. A lean, well-trained athlete carries 80,000 to 100,000 kilocalories in adipose tissue, enough fuel in theory for dozens of marathons. The constraint is not availability. Fat oxidation is too slow for race pace because the rate at which fat can be converted to ATP is biologically capped at high exercise intensities in a way that carbohydrate oxidation is not.
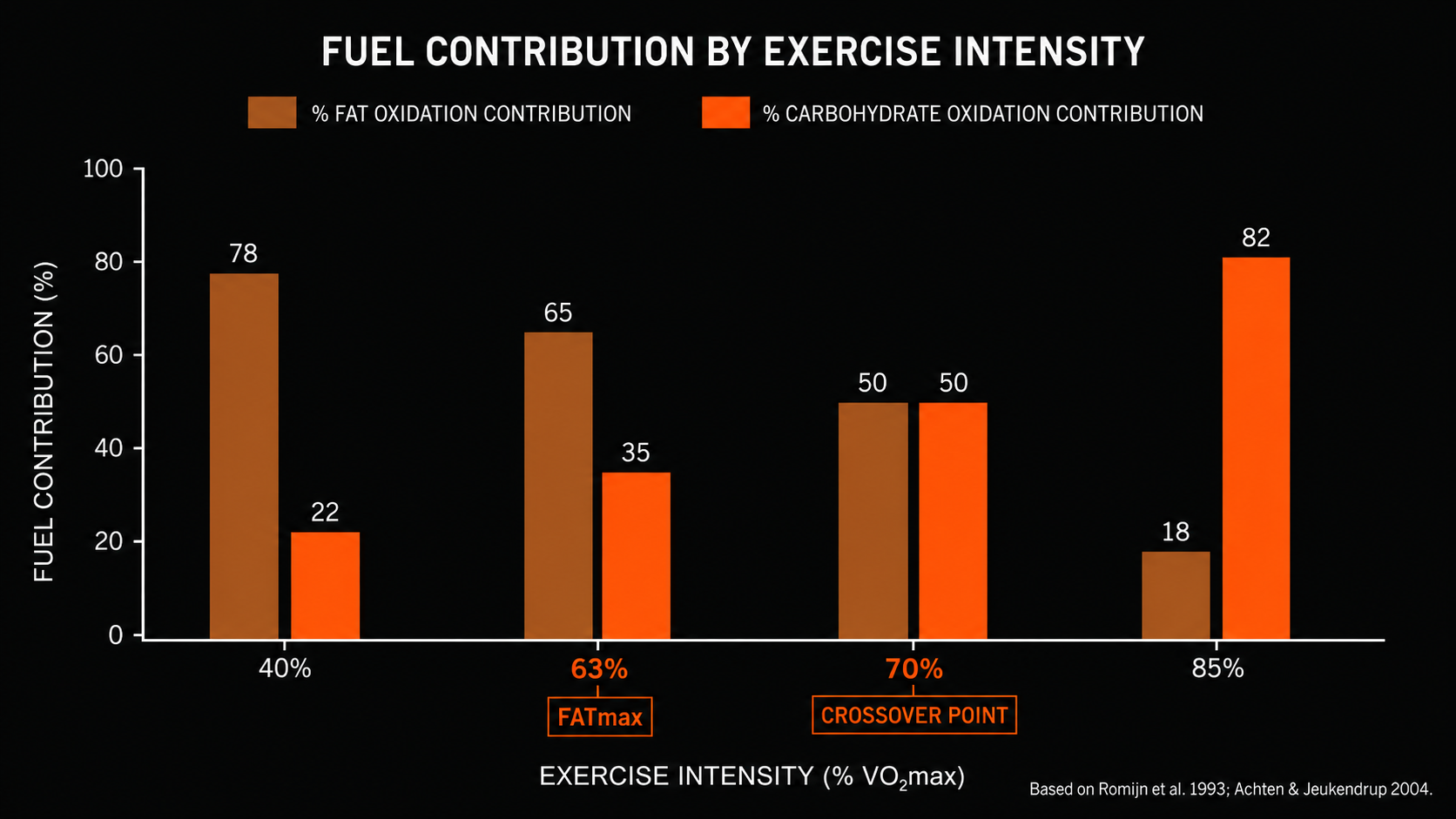
Fat oxidation requires more oxygen per unit of ATP produced than carbohydrate oxidation does. Beta-oxidation of fatty acids, and the subsequent entry of acetyl-CoA into the Krebs cycle, is metabolically more expensive in terms of oxygen demand. As exercise intensity rises and oxygen delivery becomes a limiting factor, the body preferentially uses carbohydrate because it generates more ATP per unit of oxygen consumed. This is a direct consequence of biochemistry, not a choice the metabolic system makes. The result is what researchers describe as the metabolic crossover point in exercise: the intensity at which carbohydrate oxidation surpasses fat oxidation as the dominant fuel contributor (Brooks and Mercier 1994).
Below the crossover point, fat provides a meaningful proportion of energy and fat oxidation is near its maximum rate. Above it, carbohydrate rapidly becomes the primary fuel, and fat oxidation begins to decline even as total energy demand climbs. Research on FATmax, the intensity at which fat oxidation peaks, shows that in trained endurance athletes this point occurs around 62 to 65 percent of VO2max (Achten and Jeukendrup 2004). Marathon race pace for most runners sits 10 to 20 percentage points above that ceiling.
FUEL CONTRIBUTION BY EXERCISE INTENSITY
% of total energy from each substrate
Fat oxidation %
Carbohydrate oxidation %
40% VO2max
Fat dominant
63% VO2max
FATmax
70% VO2max
Crossover
85% VO2max
Race Pace
Based on Romijn et al. 1993; Achten & Jeukendrup 2004
This is why the instruction to burn fat is an incomplete prescription when glycogen depletes. The body burns fat throughout the entire run, without interruption. The problem is that as glycogen stores fall and metabolic demand remains constant, fat cannot accelerate to cover the deficit. The crossover point does shift modestly with training: well-adapted aerobic athletes can sustain a higher fat contribution at a given pace than untrained athletes. The shift is incremental, however, and the ceiling on fat oxidation rate at marathon intensity remains intact regardless of metabolic conditioning. When glycogen falls below the rate needed to maintain workload, the result is a drop in output, not a seamless fuel transition.
Fat oxidation is too slow for race pace because the rate at which fat can be converted to ATP is biologically capped at high exercise intensities in a way that carbohydrate oxidation is not.
When glycogen falls below the rate needed to maintain workload, the result is a drop in output, not a seamless fuel transition.
Liver Glycogen, Blood Sugar, and the Brain

The muscle glycogen story accounts for the peripheral component of hitting the wall: the legs stop responding, pace becomes unsustainable, the mechanical output of working muscle degrades. The wall also produces cognitive symptoms that are separate from muscular failure: difficulty concentrating, a sense of despair or disorientation, perceived exertion that escalates beyond what the physical effort seems to warrant. These symptoms originate from a second mechanism operating in parallel.
Hepatic Glycogen Depletion During Exercise
Muscle glycogen and liver glycogen are distinct pools with distinct functions. Muscle glycogen is used directly by the contracting fibers as local fuel. Liver glycogen is broken down and released as glucose into the bloodstream, where it maintains blood sugar levels for the brain and other glucose-dependent tissues. These two pools deplete at different rates, influenced by different variables.
During prolonged exercise, the liver progressively empties its glycogen stores to sustain blood glucose output. Exogenous carbohydrate consumed during exercise directly reduces the rate of hepatic glycogen depletion during exercise by providing an external source of blood glucose maintenance (Gonzalez et al. 2016). In athletes who race without fueling, or who fuel inadequately, hepatic glycogen depletion during exercise becomes a real risk at marathon distance. When liver stores fall critically low, the liver can no longer maintain blood glucose at the rate the brain requires.
MUSCLE GLYCOGEN
Local Fuel for Working Fibers
Used directly by contracting muscle fibers. Cannot be exported to the bloodstream or shared with other muscles or the brain.
LIVER GLYCOGEN
Blood Glucose Maintenance
Broken down and released as glucose into the bloodstream to maintain blood sugar levels for the brain and other glucose-dependent tissues.
Why Blood Sugar Drops During Long Runs and What It Does to the Brain
The brain is functionally dependent on glucose during sustained high-intensity exercise. When blood sugar drops during long runs, brain function degrades in measurable ways: perceived exertion increases beyond what peripheral markers justify, concentration lapses, and the motivational signal to sustain effort weakens (Nybo and Secher 2004). This is central fatigue, meaning the fatigue originates in the central nervous system rather than in the muscle itself.
Research on blood glucose and central fatigue shows that even moderate blood glucose decline, which does not require reaching the threshold of full clinical hypoglycemia, is sufficient to alter central motor drive and perceived exertion significantly (Davis and Bailey 1997). The runner at mile 22 who describes feeling unable to continue even though their legs feel fine is often experiencing this dissociation: peripheral muscle still has fuel remaining, but blood glucose decline has compromised the central signal to use it.
Muscular glycogen depletion and hepatic glycogen-driven blood glucose decline do not always peak simultaneously. When they converge, the combined effect produces the characteristic totality of the wall, where both the body's output capacity and the brain's willingness to demand output fail at the same time.
Liver glycogen is broken down and released as glucose into the bloodstream, where it maintains blood sugar levels for the brain and other glucose-dependent tissues.
Research on blood glucose and central fatigue shows that even moderate blood glucose decline, which does not require reaching the threshold of full clinical hypoglycemia, is sufficient to alter central motor drive and perceived exertion significantly.
Why the Wall Hits Suddenly: The Threshold Effect

The wall's defining characteristic is that it does not announce itself gradually. Runners describe feeling competitive and capable at mile 19, then experiencing categorical failure two miles later. This abruptness follows directly from the non-linear relationship between glycogen availability and performance output described in the previous sections.
Performance output tracks glycogen availability reasonably well until glycogen falls below the threshold at which it can sustain current workload, then output drops sharply rather than continuing to decline proportionally. The athlete does not feel progressively worse as glycogen depletes; they feel functional until the threshold is crossed, at which point ATP supply falls short of demand in a way that cannot be compensated. The abrupt sensation of failure reflects the physics of a threshold event, not a sudden physical injury.
The blood glucose mechanism compounds this effect. As liver glycogen depletes and blood glucose drops, central fatigue amplifies the muscular failure. The two curves converge roughly simultaneously in most marathon scenarios without adequate mid-race fueling, producing a compound failure that feels more sudden than either mechanism alone would produce. This is why some runners who fuel during a race still hit the wall, and why the onset timing can shift with preparation but the underlying mechanism cannot be eliminated through fueling alone.
Performance output tracks glycogen availability reasonably well until glycogen falls below the threshold at which it can sustain current workload, then output drops sharply rather than continuing to decline proportionally.
Is Hitting the Wall Mental or Physical? The Costly Misconception

The framing of the wall as a test of mental toughness persists because it contains a kernel of truth. Pacing decisions are partly psychological. Athletes who go out at unsustainable pace in the first half of a marathon accelerate glycogen depletion and move the threshold event earlier in the race. Mental discipline in pacing changes the timing of the wall. This is where psychological factors genuinely interact with the underlying physiology.
The problem arises when this kernel of truth is extended into a claim that willpower can overcome the wall once it has begun. At the moment when muscle glycogen has fallen below the threshold required to sustain current workload, and blood glucose has dropped enough to compromise central fatigue resistance, no psychological intervention can re-synthesize substrate. The wall is a substrate accounting problem. ATP demand exceeds ATP supply, and motivational state does not appear as a variable in that equation.
BEFORE THE WALL
What Mental Discipline Changes
Pacing decisions in the first half of the race affect the rate of glycogen depletion. Mental discipline here delays the timing of the threshold event.
AFTER THE WALL BEGINS
What It Cannot Change
Willpower cannot re-synthesize glycogen. ATP demand exceeds ATP supply regardless of motivational state.
The practical cost of the mental framing is measurable: athletes who believe they can push through the wall respond to its onset by increasing effort. Higher effort increases the rate of glycogen oxidation and accelerates blood glucose decline, worsening both mechanisms simultaneously. Athletes who push harder into the wall on the grounds that it is a mental test tend to deteriorate more sharply than those who reduce pace, because reducing pace at least decreases the rate at which remaining fuel is consumed. Hitting the wall is not a failure of discipline. It is a failure of substrate supply, and understanding this distinction determines whether a runner's response to the onset of the wall makes the outcome better or worse.
ATP demand exceeds ATP supply, and motivational state does not appear as a variable in that equation.
How to Delay Hitting the Wall in a Marathon: What the Mechanism Tells You
The cellular mechanism of the wall points directly to what can shift it. The wall is a function of glycogen store size, rate of glycogen depletion, fat oxidation capacity, and blood glucose maintenance. Each of these variables responds to training and fueling in ways the mechanism predicts.
Aerobic training increases mitochondrial density in skeletal muscle. More mitochondria per fiber means greater capacity for fat oxidation at a given exercise intensity, which raises the crossover point so that trained athletes can sustain a higher fat contribution at race pace than untrained athletes (Holloszy 1967). The practical outcome is that the threshold event occurs later in the race. The mechanism is specific: it is the consequence of having more sites for beta-oxidation and electron transport chain activity per unit of working muscle tissue, not efficiency in a vague general sense.
Glycogen sparing is the observed result of this mitochondrial adaptation. Trained athletes utilize proportionally less glycogen per kilometer at marathon pace compared to less-trained athletes, because fat is covering a larger share of the energy demand. This is a biochemical consequence of improved fat oxidation capacity, not a conscious conservation strategy. Increasing the capacity of muscle cells to use oxygen in support of fat metabolism is the cellular target that training addresses, and it is one of the reasons cellular oxygen utilization is a meaningful lever for endurance athletes focused on how to delay hitting the wall in a marathon.
7.1
Mitochondrial Training
Endurance training increases mitochondrial density in skeletal muscle, raising the crossover point so trained athletes sustain a higher fat contribution at race pace. The threshold event occurs later in the race.
7.2
Glycogen Sparing
The biochemical result of improved fat oxidation capacity. Trained athletes use proportionally less glycogen per kilometer at marathon pace because fat covers a larger share of energy demand.
7.3
In-Race Fueling
Exogenous carbohydrate spares liver glycogen and maintains blood glucose. This attenuates the central fatigue component of the wall even when muscle glycogen still crosses the depletion threshold.
In-race carbohydrate fueling works through the hepatic mechanism described in the previous section. Exogenous carbohydrate does not match the rate at which muscle glycogen is being consumed at race pace; absorption and oxidation of ingested carbohydrate is too slow to substitute for muscle glycogen directly. What fueling achieves is the maintenance of blood glucose and the sparing of liver glycogen, which delays the central fatigue component of the wall. An athlete who fuels adequately during a race may reach the muscular glycogen threshold at a similar point to an unfueled athlete, but the blood glucose decline and central fatigue amplification will be attenuated. That attenuation changes the experience and outcome substantially.
One intervention that does not work: attempting to reverse the wall after it has begun by reducing pace and expecting to recover glycogen. Glycogen synthesis from blood glucose during sustained exercise is negligible. Once the threshold is crossed, the task is damage limitation. The window for effective intervention is before and during the race, not after the wall appears.
Increasing the capacity of muscle cells to use oxygen in support of fat metabolism is the cellular target that training addresses, and it is one of the reasons cellular oxygen utilization is a meaningful lever for endurance athletes focused on how to delay hitting the wall in a marathon.
The window for effective intervention is before and during the race, not after the wall appears.
Frequently Asked Questions
What does hitting the wall running feel like?
Hitting the wall in running produces a sudden, categorical collapse in pace rather than a gradual slowdown. Runners typically describe leg heaviness, cognitive fog, and an inability to maintain effort that feels disproportionate to their fitness level. These symptoms reflect simultaneous failures in muscular ATP supply and blood glucose maintenance, both of which degrade at a threshold rather than linearly.
Is hitting the wall mental or physical?
The wall is primarily a physical event driven by glycogen depletion and blood glucose decline. Mental discipline affects the timing of the wall by influencing early-race pacing decisions, but willpower cannot re-synthesize glycogen once depletion reaches the performance threshold. Pushing harder after the wall begins accelerates glycogen oxidation and worsens blood glucose decline rather than reversing either mechanism.
At what mile do most runners hit the wall in a marathon?
Most runners hit the wall between miles 18 and 22, corresponding to approximately 90 to 120 minutes of sustained effort at race pace. At that intensity, muscle glycogen stored at roughly 400 to 500 grams total falls below the threshold required to maintain current workload, producing the characteristic performance collapse regardless of fitness level or mental state.
Can you recover from hitting the wall during a race?
Recovery mid-race is not physiologically feasible. Glycogen synthesis from blood glucose during sustained exercise is negligible, meaning depleted stores cannot be meaningfully rebuilt while running. Slowing pace reduces the rate of further glycogen depletion and limits blood glucose decline, but does not restore the performance capacity that existed before the threshold was crossed.
What is the difference between hitting the wall and bonking?
The terms are often used interchangeably, but bonking more specifically refers to hypoglycemia, a significant drop in blood glucose, as the primary mechanism of collapse. Hitting the wall encompasses both muscular glycogen depletion and blood glucose decline. In practice, the two mechanisms occur together during a marathon, and the symptoms of each compound the other.
The Bottom Line
The wall is a substrate event. Muscle glycogen depletes below the threshold at which it can supply ATP fast enough to sustain race pace. Fat oxidation, rate-limited by beta-oxidation kinetics and oxygen cost, cannot accelerate to cover the deficit. Liver glycogen drains to the point where blood glucose drops, and central fatigue compounds the peripheral failure. These mechanisms converge at a predictable point in the marathon, not because of poor mental preparation, but because of the finite size of the glycogen tank relative to the metabolic demand of sustained high-intensity aerobic work.
The insight this produces is not that the wall is inevitable. It is that the strategies worth investing in are the ones that address the actual variables: increasing mitochondrial capacity to shift the crossover point, maintaining fueling to protect blood glucose, and calibrating pace to match the rate of glycogen depletion to the distance remaining. An athlete who understands the cellular mechanism of the wall is in a fundamentally different position than one who interprets it as a character test, not because understanding changes what happens biochemically, but because it determines whether the response before, during, and at the onset of the wall works with the physiology or against it.
An athlete who understands the cellular mechanism of the wall is in a fundamentally different position than one who interprets it as a character test, not because understanding changes what happens biochemically, but because it determines whether the response before, during, and at the onset of the wall works with the physiology or against it.
The wall is not a test of will. It is a threshold event in substrate availability — and athletes who understand that treat it as a cellular problem, which means they prepare for it instead of trying to push through it.
Glycogen has a hard ceiling, fat oxidation has a rate ceiling, and blood glucose maintenance has its own tank. The wall is what happens when all three converge. Preparation changes when they converge. It does not change whether they can.
Mydos Performance
— Cellular Energy & Fuel Systems Series —
The Physiology of Hitting the Wall Running
Glycogen, Fat Oxidation, and Why the Marathon Threshold Arrives Without Warning
· By Ricardo Londono, MD/PhD ·