
The Energy Assumption That Gets High-Performing Athletes Wrong

The three energy systems of the body are the actual mechanism behind every physical and cognitive output. When a serious athlete experiences persistent low energy or a performance plateau despite consistent training, the instinct is to search for the behavioral variable that changed: stress level, sleep quality, nutritional consistency, accumulated fatigue. These factors matter, but they operate upstream of the fundamental constraint. The body cannot convert sleep directly into muscular force, and food does not power muscle contraction without an intermediate step. Every substrate consumed, every night recovered, only translates to physical output if the cell can convert available material into adenosine triphosphate.
Energy in body systems is governed by biochemical rules, not motivational ones. A 40-year-old athlete experiencing declining output is not necessarily doing something wrong at the behavioral level. The more precise diagnostic question is whether the cell's ATP production rate is adequate for the demand being placed on it, and if not, which biological pathway is the limiting constraint. This is a specific, addressable question. The answer runs through three enzymatic systems that operate according to rules that do not change with willpower or schedule. The foundational account of how those three systems are structured and interact — including why all three run at once rather than in sequence — is in the three energy systems explained.
Energy in body systems is governed by biochemical rules, not motivational ones.
How Cells Produce Energy: ATP and the One Currency That Powers Everything
ATP is the only form of energy the cell's machinery can directly use. Glucose, fat, carbohydrate, and creatine phosphate do not power muscle contraction or nerve firing directly. These substrates are inputs that cellular pathways convert into ATP. The actual work of physiology is performed by the molecule that results from that conversion.
The human body contains approximately 250 grams of ATP at any given moment (Hargreaves et al. 2020). At maximal physical effort, this supply is depleted within a few seconds. The body does not store large quantities of ATP because the molecule is metabolically active and costly to maintain in reserve. Instead, three enzymatic pathways continuously resynthesize ATP during activity, converting available substrates fast enough to meet current demand. When resynthesis rate falls behind demand rate, performance degrades at a pace proportional to the gap.
250g
Total Body ATP at Rest
<10 sec
Time to Depletion at Max Effort
The cell's operational problem is therefore not whether enough fuel exists. A typical trained athlete carries enough stored fat to fuel dozens of hours of low-intensity effort. The limiting variable is whether the cellular machinery can convert that fuel into ATP at the rate the current workload requires. That conversion happens through three distinct pathways, each operating at a different speed and drawing on different substrates.
The limiting variable is whether the cellular machinery can convert that fuel into ATP at the rate the current workload requires.
The Three Energy Systems of the Body: Speed, Capacity, and Trade-offs
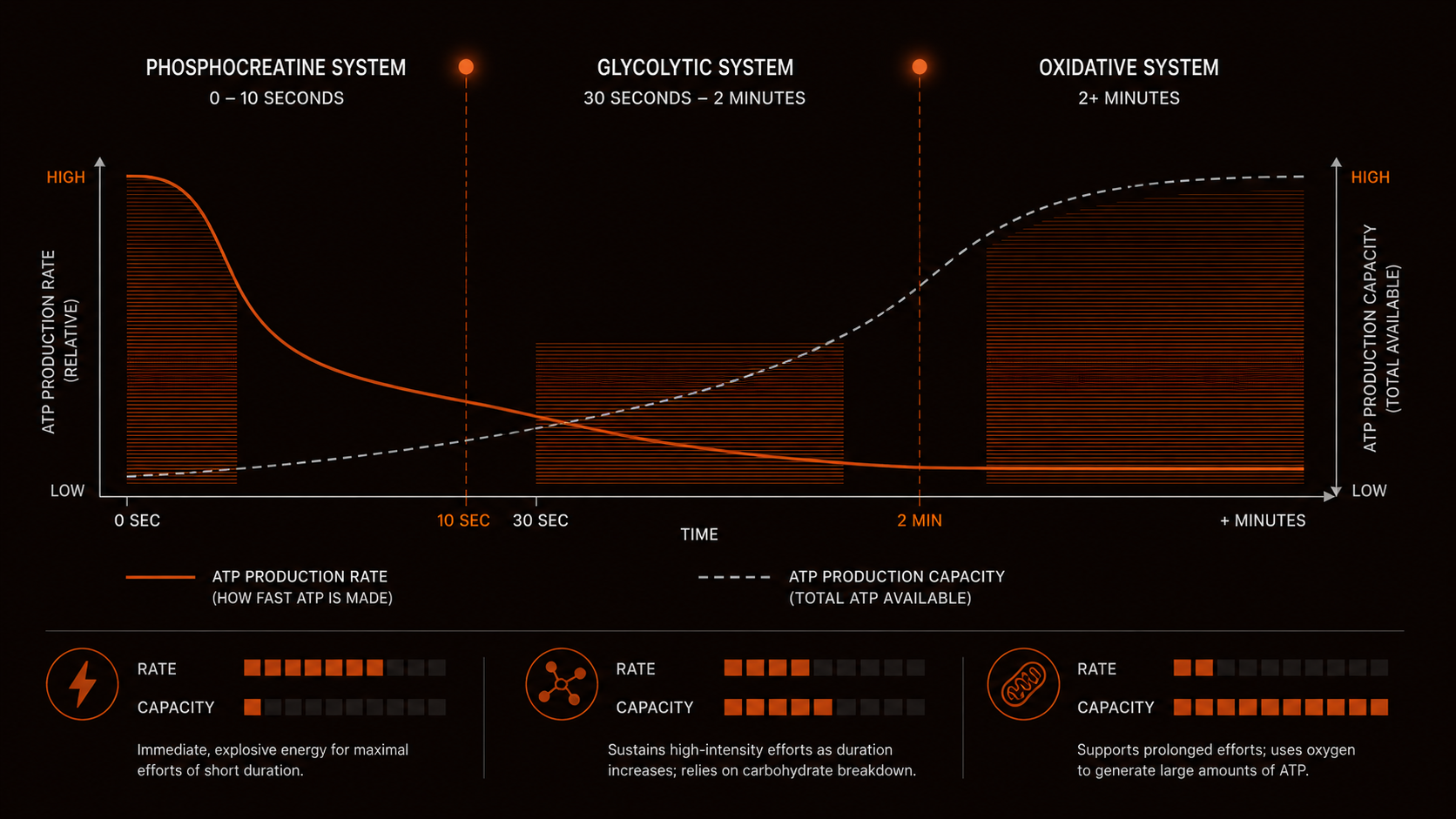
The three energy systems of the body resynthesize ATP through different biochemical mechanisms at different rates. At any given moment, all 3 energy systems in the body are active, with the relative contribution of each pathway shifting based on exercise intensity, duration, and available substrates. They do not activate sequentially. They operate simultaneously, and the dominant pathway at any moment is the one best matched to current ATP demand.
Three ATP Pathways
Phosphocreatine System
Maximal power, 6–10 seconds. Requires no oxygen. Limited by creatine phosphate stores.
Glycolytic System
Fast ATP, 30 sec–3 min. No oxygen required. Limited by hydrogen ion and inorganic phosphate accumulation.
Oxidative System
Sustained output, unlimited duration. Requires oxygen and mitochondria. Limited by mitochondrial capacity.
The Phosphocreatine System
The phosphocreatine system is the fastest ATP resynthesis pathway available to the cell. A creatine phosphate molecule donates its phosphate group directly to ADP, reforming ATP in a single enzymatic step that requires no oxygen and produces no fatiguing byproducts in the immediate process. This makes the phosphocreatine system the default energy source for any maximal-intensity effort: the first seconds of a sprint, the initial pull of a heavy barbell, the opening exchange of a sparring round. The full cellular account of what happens at these intensities — including how the anaerobic energy system governs peak output — is in the adjacent article in this pillar.
The system's limitation is capacity, not speed. Intramuscular creatine phosphate stores are depleted within six to ten seconds of maximal effort, and full restoration takes approximately three to five minutes of aerobic recovery, with the resynthesis rate driven primarily by the oxidative system (Bogdanis et al. 1996). This is the mechanistic basis for long rest intervals in power training: the objective is full PCr availability for each effort, not an accelerated recovery window that reduces the contribution of the system being targeted.
The Glycolytic System
The glycolytic system resynthesizes ATP by breaking down glucose through a series of enzymatic reactions in the cell cytoplasm, with no oxygen required. The process is faster than oxidative metabolism, making it the dominant energy source for sustained high-intensity efforts lasting from roughly thirty seconds to two to three minutes. A hard grappling exchange, a conditioning circuit, or an extended sprint depends substantially on glycolytic ATP production.
The fatigue from sustained glycolytic effort is an acidosis and inorganic phosphate accumulation problem, not a lactate toxicity problem.
The byproducts of this process, primarily hydrogen ions and inorganic phosphate, accumulate as glycolytic rate increases. This accumulation interferes with further enzymatic activity and impairs muscle contractile function, producing the acute fatigue and declining power output that characterize hard glycolytic work. Lactate accumulates simultaneously but functions primarily as a fuel: it is exported to adjacent cells and oxidized there, a process described by the lactate shuttle framework (Brooks 2018). The fatigue from sustained glycolytic effort is an acidosis and inorganic phosphate accumulation problem, not a lactate toxicity problem (Westerblad et al. 2002).
The Oxidative System
The oxidative system resynthesizes ATP through complete oxidation of glucose and fat in the mitochondria via the Krebs cycle and the electron transport chain. Of the three pathways, it is the slowest in terms of ATP production rate per unit time, but it produces by far the most ATP per unit of substrate: approximately 30 molecules from complete glucose oxidation, compared to 2 from anaerobic glycolysis (Hinkle et al. 2005). Fat oxidation yields even more ATP per molecule, making the oxidative system the basis for all sustained efforts beyond a few minutes and the primary driver of recovery between intense efforts.
The oxidative system's practical ceiling is not substrate availability but mitochondrial capacity. A muscle cell may have fat and carbohydrate available in abundance, but it can only oxidize those substrates as fast as its mitochondria allow. Mitochondrial number, density, and functional efficiency collectively determine how much ATP the oxidative pathway can produce per unit time. When mitochondrial capacity declines, the oxidative system bottlenecks at lower work rates, the glycolytic system compensates earlier than it should, and fatigue accumulates at intensities where it previously did not. The variable that determines oxidative capacity is mitochondrial biogenesis.
When mitochondrial capacity declines, the oxidative system bottlenecks at lower work rates, the glycolytic system compensates earlier than it should, and fatigue accumulates at intensities where it previously did not.
What Is Mitochondrial Biogenesis and Why Your Oxidative Capacity Depends on It

The ceiling of the oxidative system is not substrate availability but mitochondrial capacity, and that capacity is determined by a regulated biological process called mitochondrial biogenesis. It is not a continuous background process but a stimulus-driven response to specific physiological conditions, primarily sustained aerobic effort and the metabolic signals that effort generates.
The central regulator is PGC-1alpha (peroxisome proliferator-activated receptor gamma coactivator 1-alpha). Exercise activates PGC-1alpha signaling in skeletal muscle, and PGC-1alpha drives the transcription of genes involved in producing new mitochondria and increasing mitochondrial enzyme density (Baar et al. 2002). The training stimulus that most durably upregulates this pathway is not maximal or near-maximal intensity. Sustained low-to-moderate aerobic work generates the specific metabolic conditions, including shifts in the AMP:ATP and NAD+:NADH ratios, that activate PGC-1alpha signaling most effectively.
A muscle with higher mitochondrial density sustains greater aerobic output before the glycolytic system is recruited. Conversely, when mitochondrial density declines, the oxidative threshold falls: the glycolytic system compensates at lower intensities, hydrogen ions and inorganic phosphate accumulate faster, and perceived effort at a given workload increases. The practical experience is a training load that previously felt manageable now requiring more effort than the work rate warrants, with slower recovery between rounds and a lower ceiling on sustained output.
High Mitochondrial Density
Sustained aerobic output
Glycolytic system is recruited only at genuinely high intensities. Recovery is faster. Perceived effort at moderate workloads remains low.
Low Mitochondrial Density
Early glycolytic compensation
Glycolytic system activates at moderate intensities. Hydrogen ion accumulation begins sooner. Perceived effort rises faster at the same workload.
Mitochondrial function declines measurably with age. Research on human skeletal muscle in vivo documents a significant decrease in mitochondrial ATP production rate and mitochondrial abundance from middle age onward, driven by reduced biogenesis signaling and accumulated mitochondrial damage (Short et al. 2005). This decline occurs independent of training effort, though appropriate training substantially attenuates it. An athlete training with the same frequency and intent at 45 as at 30 may produce meaningfully less cellular output, not because effort has changed, but because the mitochondrial system generating that output has degraded at a rate that behavioral consistency alone does not counteract. The other determinant of how well the oxidative system performs is whether the cell selects the appropriate fuel at the appropriate intensity, a capacity called metabolic flexibility.
An athlete training with the same frequency and intent at 45 as at 30 may produce meaningfully less cellular output, not because effort has changed, but because the mitochondrial system generating that output has degraded at a rate that behavioral consistency alone does not counteract.
What Is Metabolic Flexibility: Why Fuel Switching Determines Sustained Output

Fat and carbohydrate are not used in equal proportion across all exercise intensities. The cell selects between them based on intensity, substrate concentration, and the functional state of the mitochondria. Metabolic flexibility describes how efficiently a cell executes this selection: at low-to-moderate intensities, a flexible cell burns predominantly fat; as intensity rises and fast-twitch fibers are recruited, it shifts toward carbohydrate; and it transitions smoothly between these states without creating a mismatch between substrate use and available supply.
The physiological signals governing fuel selection include insulin levels, substrate concentrations in blood and muscle, and mitochondrial oxidative capacity. The last factor is the most consequential for performance. Fat oxidation is an exclusively aerobic process that occurs entirely within the mitochondria. When mitochondrial function is impaired, the rate of fat oxidation falls, and the cell relies more heavily on carbohydrate even at intensities where fat oxidation should dominate. This is the cellular mechanism of metabolic inflexibility, and it is not primarily a dietary problem (Goodpaster and Sparks 2017).
An athlete with poor metabolic flexibility depletes glycogen stores prematurely because carbohydrate is consumed at intensities a metabolically flexible athlete handles largely on fat. The glycolytic system operates at higher rates than the work rate demands, accumulating byproducts and producing early fatigue at outputs that should be sustainable. Correcting this requires improving mitochondrial oxidative capacity, not adjusting macronutrient ratios. The machinery processes the fuel. Changing the fuel without fixing the machinery shifts which substrate runs out first, not whether the system can meet demand.
Metabolically Flexible
Fat fuels the base, glycogen fuels the peak
At moderate intensities, fat oxidation provides the majority of ATP. Glycogen is preserved for high-intensity demands and depletes slowly.
Metabolically Inflexible
Glycogen consumed at all intensities
The cell defaults to carbohydrate at moderate intensities. Glycogen depletes prematurely. Fatigue accumulates earlier than intensity warrants.
Aging and chronic training that neglects low-intensity aerobic work both reduce metabolic flexibility through the same mechanism: progressive decline in mitochondrial oxidative capacity. An athlete who has trained predominantly at high intensities for years may have a well-developed glycolytic system and a comparatively underdeveloped oxidative pathway, leaving the cell metabolically inflexible regardless of dietary quality.
This is the cellular mechanism of metabolic inflexibility, and it is not primarily a dietary problem.
What Cellular Energy Failure Actually Looks Like in Training and Performance

Cellular energy deficits do not always present as generalized fatigue. They present as specific, repeating patterns that do not resolve with additional rest or adjusted nutrition. The most common is a premature shift to glycolytic metabolism at intensities that should be handled aerobically. One of the clearest expressions of this shift is what happens when effort extends deep into glycolytic territory — a pattern examined in detail in the article on hitting the wall running.
6.1
Premature Glycolytic Shift
Burning and high perceived effort at moderate intensities. Recovery between rounds slower than effort warrants. Caused by insufficient oxidative capacity.
6.2
Cognitive Decline Under Load
Difficulty maintaining focus during training. Post-session brain fog. Afternoon mental fatigue following intense morning work.
6.3
Performance Plateau Without Overtraining
Consistent training, no adaptation. More volume doesn't break the ceiling. The oxidative system is the bottleneck, not training effort.
An athlete with limited oxidative capacity experiences a faster-than-expected rise in perceived effort at moderate intensity, a burning sensation in the muscles at work rates that previously felt manageable, and a recovery rate between rounds or sets that seems disproportionate to the effort expended. These patterns occur because the glycolytic system is recruited earlier than the intensity warrants, accumulating hydrogen ions and inorganic phosphate at rates that appropriate aerobic output would not generate. The cause is insufficient mitochondrial capacity to sustain the oxidative pathway at the required output rate, not poor conditioning in the conventional sense.
Cognitive function follows the same cellular logic. The brain competes for ATP from the same supply as working muscle and does not have the substrate reserves of skeletal tissue. When overall ATP production is limited, CNS output is one of the first things affected. Difficulty maintaining focus during training, post-session mental fatigue extending for several hours, and afternoon cognitive decline following intense morning work are consistent with an energy production system that cannot fully meet demand. These are cellular phenomena expressed in neurological output, not independent problems requiring separate fixes.
The cause is insufficient mitochondrial capacity to sustain the oxidative pathway at the required output rate, not poor conditioning in the conventional sense.
The pattern that most clearly points to a cellular constraint rather than a behavioral one is a performance plateau that persists despite consistent, adequate training. An athlete managing all behavioral variables correctly, who continues to underperform relative to training input, is encountering a ceiling defined by cellular capacity. Applying more volume to an insufficient oxidative foundation does not raise that ceiling. It increases demand on the system that is already the bottleneck. The most common response to these patterns is to apply a behavioral intervention to what is a cellular problem, which is why the symptoms persist.
These are cellular phenomena expressed in neurological output, not independent problems requiring separate fixes.
The Wrong Diagnosis: Why Treating Output Problems as Input Problems Doesn't Work

Behavioral interventions target the inputs to the energy system: fuel availability, recovery duration, arousal state. They do not address the efficiency of the machinery converting those inputs into ATP. A behavioral fix applied to a cellular problem can reduce the perception of constraint without resolving its source.
7.1
Caffeine
Blocks adenosine receptors and reduces perceived fatigue. Does not increase mitochondrial ATP production or expand oxidative capacity. Masks the constraint without resolving it.
7.2
More Training Volume
Applying additional demand to an already-bottlenecked oxidative system produces no adaptation or regression. The stimulus for biogenesis is specific, not maximal.
7.3
Substrate Optimization
Adjusting carbohydrate or fat intake changes which fuel is available to the three energy systems. It does not change the mitochondrial machinery that processes those fuels.
Caffeine is the most common example. Adenosine accumulates as a byproduct of ATP hydrolysis during sustained effort and binds to receptors in the brain, increasing perceived exertion. Caffeine blocks those receptors, reducing perceived effort at a given workload and extending time to task failure. This is a genuine, measurable effect on performance. What caffeine does not do is increase mitochondrial ATP production, accelerate PCr resynthesis, or expand the oxidative pathway's capacity. It changes the perceived level of the cellular constraint without changing the constraint itself.
Adding training volume in response to a plateau follows the same failed logic. Applying greater demand to an oxidative system that is already the bottleneck produces either no adaptation or regression: the stimulus for mitochondrial biogenesis is not maximal metabolic stress but specific, appropriately dosed aerobic work. The correct response to an oxidative deficit is a different distribution of training intensity, not more volume at the same intensities that failed to produce adaptation previously. Patterns of accumulated effort without recovery — and the consequences — are examined in overtraining syndrome in athletes.
Nutritional strategies including carbohydrate periodization, fat adaptation protocols, and ketogenic approaches modify the fuel inputs available to the three energy systems without changing the mitochondrial machinery that processes those fuels. A well-fueled cell with low mitochondrial density still produces less ATP per unit time than a cell with high mitochondrial density. Substrate availability is rarely the primary limiter for an athlete consuming adequate calories. Treating it as the lever produces selective substrate optimization while the underlying capacity constraint remains.
A behavioral fix applied to a cellular problem can reduce the perception of constraint without resolving its source.
What a Metabolic Flexibility Training Program Actually Does at the Cellular Level

The cellular mechanism by which training improves energy output is biogenesis, not substrate manipulation. Understanding this is what a metabolic flexibility training program is actually designed to accomplish at the molecular level: expanding the mitochondrial capacity of the oxidative pathway so that fat oxidation can sustain a wider range of intensities before glycolytic compensation is required.
Sustained aerobic exercise at low-to-moderate intensity generates changes in the cell's metabolic environment, specifically shifts in the AMP:ATP and NAD+:NADH ratios, that activate PGC-1alpha signaling. PGC-1alpha drives the transcription of genes responsible for producing new mitochondria and increasing the density of oxidative enzymes (Egan and Zierath 2013). This is the mechanism documented in foundational exercise physiology research (Holloszy 1967) and refined substantially since: aerobic training expands oxidative capacity by increasing the number and functional density of the organelles that execute the oxidative pathway.
High-intensity training produces different adaptations. Near-maximal and supramaximal efforts primarily stress the glycolytic and phosphocreatine systems, producing fitness gains in those pathways through different signaling cascades. These adaptations matter for power output and high-intensity endurance, but they do not substantially increase mitochondrial mass or improve fat oxidation capacity. An athlete training exclusively at high intensity accumulates glycolytic fitness without building the aerobic foundation that determines how quickly the glycolytic system recovers between efforts and how long high-intensity work can be sustained over a training session.
Low-Intensity Aerobic Training
PGC-1alpha activation → mitochondrial biogenesis
Shifts AMP:ATP and NAD+:NADH ratios, activating PGC-1alpha. Drives production of new mitochondria. Expands fat oxidation capacity and raises the intensity threshold before glycolytic compensation begins.
High-Intensity Only Training
Glycolytic adaptation without aerobic base
Develops the phosphocreatine and glycolytic pathways. Does not substantially increase mitochondrial mass. Produces glycolytic fitness without expanding the oxidative foundation that determines how well that fitness can be sustained and recovered.
The practical inference is that improving cellular energy output requires a training intensity distribution that allocates meaningful volume to the oxidative pathway specifically. This is the mechanistic basis for the polarized training structures that appear consistently in performance research: a large proportion of aerobic volume at intensities where the oxidative system is the primary ATP source, combined with a smaller volume of high-intensity work that develops the power and glycolytic ceiling the aerobic base supports. The framework for structuring this distribution across the training cycle is covered in the fitness-fatigue model. The cellular machinery is trainable, and in some cases, specific compounds can also support the same molecular pathways that training targets. What the relationship between aerobic conditioning and oxygen delivery capacity means for how these training intensities translate to performance is covered separately.
The cellular mechanism by which training improves energy output is biogenesis, not substrate manipulation.
Frequently Asked Questions
What are the three energy systems of the body?
The three energy systems are the phosphocreatine system (maximal, 6–10 seconds), the glycolytic system (high-intensity, up to 2–3 minutes), and the oxidative system (sustained aerobic effort). They all operate simultaneously, with the dominant pathway shifting based on exercise intensity and duration. All three resynthesize ATP, the only energy currency cells can directly use.
Why does energy decline with age even in athletes who train consistently?
Mitochondrial biogenesis rate slows with age, and existing mitochondria accumulate damage over time. This reduces the oxidative system's ATP production capacity, forcing earlier reliance on the glycolytic system at lower work rates. The result is faster fatigue, slower recovery, and a performance ceiling that consistent training effort alone does not raise.
What is metabolic flexibility and why does it matter for performance?
Metabolic flexibility is the cell's capacity to shift between fat and carbohydrate oxidation based on exercise intensity. An athlete with high metabolic flexibility burns predominantly fat at low-to-moderate intensities, conserving glycogen for high-intensity demands. Poor flexibility forces carbohydrate use at all intensities, depleting glycogen prematurely and generating more fatigue byproducts at moderate output.
What causes fatigue during high-intensity exercise?
During high-intensity glycolytic effort, fatigue is caused primarily by accumulation of hydrogen ions and inorganic phosphate in the muscle cell. These compounds interfere with the enzymatic processes of further ATP resynthesis and impair muscle contractile function. Lactate accumulates simultaneously but is a fuel, not a toxin, and is not a primary fatigue driver.
What is mitochondrial biogenesis?
Mitochondrial biogenesis is the process by which cells increase mitochondrial number and mass in response to physiological stimuli. The primary trigger is sustained low-to-moderate aerobic exercise, which activates PGC-1alpha signaling. More mitochondria means greater oxidative ATP production capacity, which translates to better endurance, faster recovery, and sustained output at higher intensities.
Cordyceps Mushroom for Energy: The Cellular Argument
Low energy and underperformance, framed correctly, are cellular phenomena. The three energy systems produce ATP at a rate that depends on the functional capacity of the mitochondrial oxidative pathway, the available substrate, and the cell's ability to select appropriately between substrates at a given intensity. When that rate is insufficient for demand, the experience is a measurable shortfall in cellular output: early fatigue, slow recovery, cognitive decline under load, and a performance ceiling that behavioral adjustments cannot raise.
This framing has a direct implication for what kinds of support are relevant. A compound that interacts with the oxidative pathway at a cellular level is operating on the mechanism that determines energy output. Cordyceps mushroom for energy production is proposed to work through this kind of interaction. The proposed mechanism involves adenosine analogue compounds in cordyceps, including cordycepin, that interact with adenosine signaling pathways involved in cellular energy metabolism. A controlled trial of Cs-4 cordyceps extract in healthy older adults found significant improvement in metabolic threshold and ventilatory threshold after twelve weeks compared to placebo, consistent with an effect on oxidative pathway function (Chen et al. 2010). The mechanistic picture is not fully characterized, and the evidence base is limited in scale. The biological target, however, is the oxidative pathway, placing it in a different category from stimulant compounds that suppress the perception of cellular constraints without addressing their source.
The more fundamental conclusion stands independent of supplementation choices. The three energy systems operate according to biochemical rules, and those rules include the progressive decline of mitochondrial biogenesis rate and oxidative capacity with age. A 40-year-old athlete who trains seriously and still underperforms is encountering a cellular phenomenon, not a motivational or behavioral one. That is a specific cause. It has specific interventions: training that targets the oxidative pathway, rebuilds mitochondrial density, and restores the cellular foundation that performance depends on at every intensity.
A 40-year-old athlete who trains seriously and still underperforms is encountering a cellular phenomenon, not a motivational or behavioral one.
Your energy is a cellular output. The three systems that produce it respond to biochemical stimuli — and when those systems decline, no amount of effort or optimization above the cellular level compensates.
Mitochondrial density, fuel-switching capacity, and ATP resynthesis rate are trainable. The interventions that move them are specific. Understanding the mechanism is the prerequisite for applying them correctly.
Mydos Performance
— Cellular Energy & Fuel Systems Series —
The Three Energy Systems of the Body: Why Low Energy and Underperformance Start at the Cellular Level
How your cells actually produce power — and why that capacity declines with age
· By Ricardo Londono, MD/PhD ·